
Jednym z przykładów nieredukowalnej złożoności jest proces, który większość z nas w chwili skaleczenia się uważa za coś oczywistego - chodzi o krzepnięcie krwi. Kiedy byłem małym chłopcem i zdarzyło mi się skaleczyć to byłem przekonany, że krew na mojej ranie krzepnie, ponieważ ma kontakt z powietrzem. Uważałem że w kontakcie z nim wysusza się i dlatego robi się strup.
Karol Darwin być może miał podobne wyjaśnienie, kiedy obserwował tworzący się skrzep, ponieważ nie miał najmniejszego pojęcia o zawiłościach biochemicznych, które towarzyszą powstawaniu skrzepu tamującego upływ krwi. Być może gdyby miał choć część tej wiedzy na swoim naukowym koncie, to jego teoria nigdy nie ujrzałaby światła dziennego!
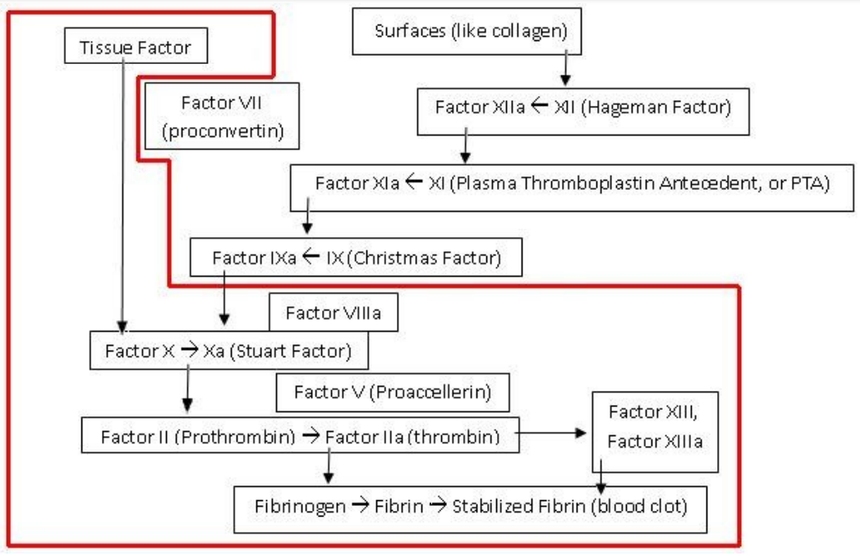
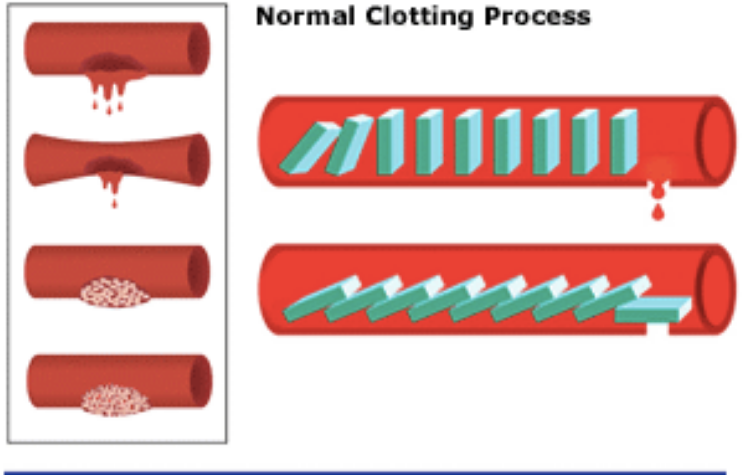
Normalnie dzieje się tak, że z przedziurawionego naczynia natychmiast zaczyna wypływać zawarty w nim płyn i cieknie dopóty, dopóki się ono nie opróżni. Jeżeli jednak przekłujemy czy przetniemy sobie skórę, upływ krwi szybko ustaje, gdyż tworzy się skrzep. Ale jak o tym dobrze wiedzą lekarze, krzepnięcie krwi jest bardzo złożonym, wielostopniowym procesem, w którym biorą udział liczne, oddziałujące na siebie białka. Uczestniczą one w tak zwanej kaskadowej aktywacji krzepnięcia. Ten delikatny proces leczniczy w ogromnej mierze zależy od tego, kiedy i z jaką szybkością zachodzą różne reakcje. Gdyby coś się źle potoczyło, człowiek mógłby się wykrwawić na śmierć albo przeciwnie, cała jego krew mogłaby ulec skrzepnięciu. Moment rozpoczęcia reakcji i ich szybkość mają więc żywotne znaczenie.
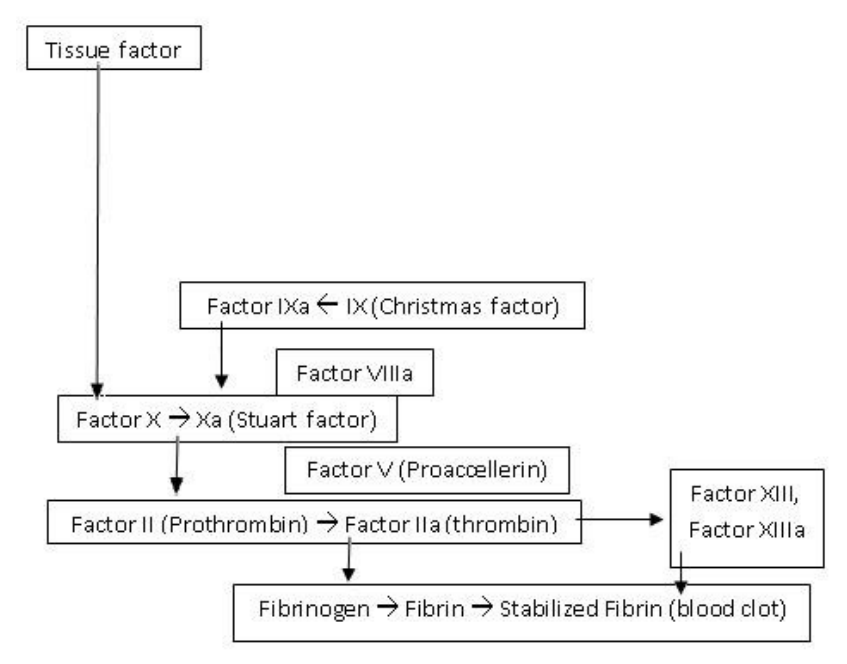
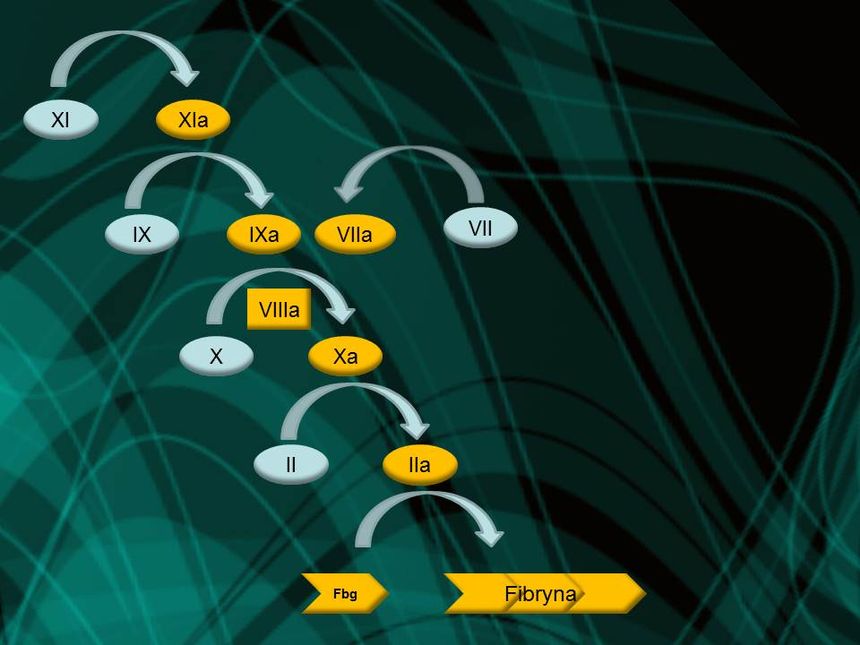
Z badań biologicznych wynika, iż w krzepnięciu krwi bierze udział wiele czynników i żadnego z nich nie może brakować. Chociaż istnieją różnice w składzie białek tworzących kaskadę krzepnięcia u pewnych gatunków, to zawsze konieczna jest minimalna ilość tych cząsteczek, która tworzy nieredukowalnie złożony rdzeń:

Biochemik Michael Behe zadaje pytanie: „Jak to się dzieje, że raz rozpoczęty proces krzepnięcia zostaje przerwany, zanim jeszcze cała krew (…) zamieni się w ciało stałe?” Wyjaśnia następnie, „iż tworzenie się skrzepu, wstrzymywanie dalszego krzepnięcia, wzmacnianie skrzepu oraz usuwanie go” stanowią zintegrowany system biologiczny. Jeżeli któryś element źle zadziała, cały system nie spełni swej funkcji.
Ewolucjonista Russell Doolittle, będący profesorem biochemii na Uniwersytecie Kalifornijskim, pyta: „Jakimże sposobem mógł powstać w wyniku ewolucji tak skomplikowany i precyzyjny proces? (…) Paradoksalna sytuacja: skoro każde białko musi zostać uaktywnione przez inne, to jak ten system mógł się rozwinąć? Jakiż pożytek przynosiła dowolna jego część, dopóki nie działała całość?” Doolittle stara się wyjaśnić pochodzenie tego procesu, posługując się ewolucyjną argumentacją. Jednakże profesor Behe zwraca uwagę, że „aby odpowiednie geny znalazły się na odpowiednich miejscach, potrzebny byłby niezwykle szczęśliwy traf”. Wskazuje także, iż wyjaśnienie podane w nonszalancki sposób przez Doolittle’a skrywa olbrzymie trudności.
Tak więc jednym z podstawowych zarzutów wobec modelu ewolucyjnego jest fakt, że nie wyjaśnia on pochodzenia tego, co prostsze być nie może, czyli nieredukowalnego rdzenia. Profesor Michael Behe oznajmia: „Chciałbym podkreślić, że dobór naturalny, będący motorem darwinowskiej ewolucji, działa jedynie wtedy, gdy istnieje coś, co może zostać wyselekcjonowane - coś, co przynosi pożytek już teraz, a nie dopiero w przyszłości”. No tak; dobór naturalny nie ma możliwości proroka :) Nawet uczciwi przeciwnicy teorii inteligentnego projektu w przyrodzie przyznają, że darwiniści nie mają pojęcia, w jaki sposób układy niekompletne, które rzekomo były ewolucyjnymi prekursorami dzisiejszych systemów biochemicznych, mogły zdobywać przewagę selekcyjną.
Michael Behe to jeden z najlepszych biologów teoretycznych [ewolucyjnych] w naszych czasach. To nie bakteriolog typu PZ Myersa, dziennikarz rodzaju Dawkinsa czy genetyk populacji w typie Jerry Coyne’a. To wytrawny znawca ewolucjonizmu molekularnego, specjalista od struktury białek i prawdziwy naukowiec. Np. tutaj jest jego rewelacyjna praca z dziedziny biologii ewolucyjnej opublikowana w porządnej literaturze naukowej;
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2286568/

Jego wnioski o nieredukowalnej złożoności systemów biologicznych nie są wynikiem chciejstwa, tylko gruntownej analizy naukowej przy zastosowaniu tego samego aparatu poznawczego (teoretycznego), którym posługują się wszyscy inni biolodzy teoretyczni. Behe’ego można spotkać na każdej znaczniejszej konferencji poświęconej biologii ewolucyjnej. Jeżeli ktoś nie rozumie dlaczego mówimy o nieredukowanych sieciach genetycznych, to proszę poczytać, co pod inną nazwą niż nieredukowalna złożoność twierdzą sami darwiniści.
Autorzy artykułu w wikipedii bardzo się starali utopić istotę konsekwencji dla neodarwinizmu – fakt powszechnego występowania nieredukowalnej złożoności w biologii molekularnej i w efekcie wyszedł im niezły bełkot. To mieszanie genetyki populacyjnej z ewolucjonizmem molekularnym przypomina mieszanie wody z olejem. Dlatego też bardzo łatwo można odcedzić istotę problemu ukrywaną w słowotoku, zaprawionym różnymi sztuczkami retorycznymi, jaką jest fakt istnienia nieredukowalnej złożoności systemów biochemicznych. Jak zwał, tak zwał: nieredukowalna złożoność, czy silna negatywna epistaza, zawsze negatywna dla tendencyjnych „wyjaśnień” od których w tekście się roi oraz dla możliwości stopniowej ewolucji złożonych z wielu nierozerwalnie zintegrowanych i dopasowanych części maszyn molekularnych ????
https://en.wikipedia.org/wiki/Epistasis

[.....]




Naukowcy wyeliminowali jeden gen i sieć genetyczna nie spełniła celu funkcji:






http://pandasthumb.org/archives/2008/05/behe-vs-lamprey.html
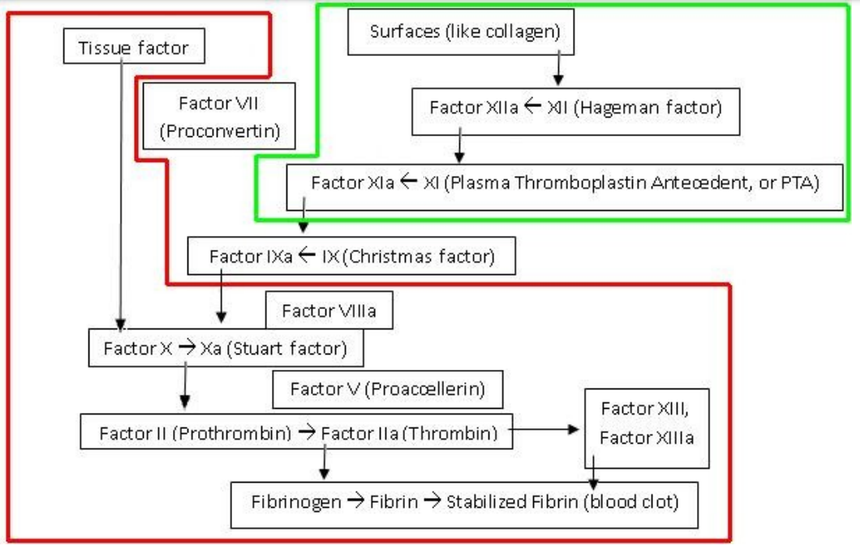
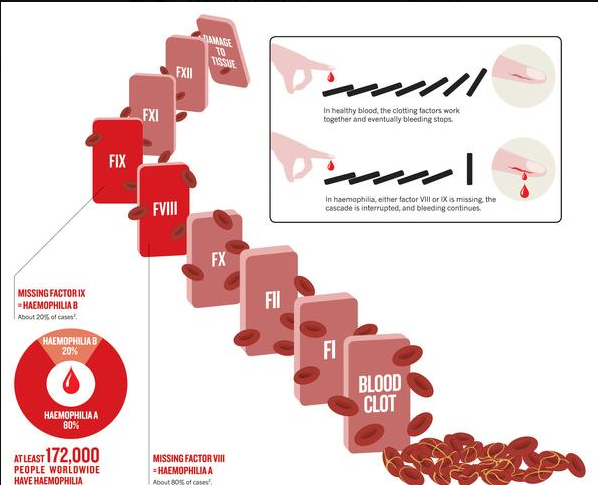
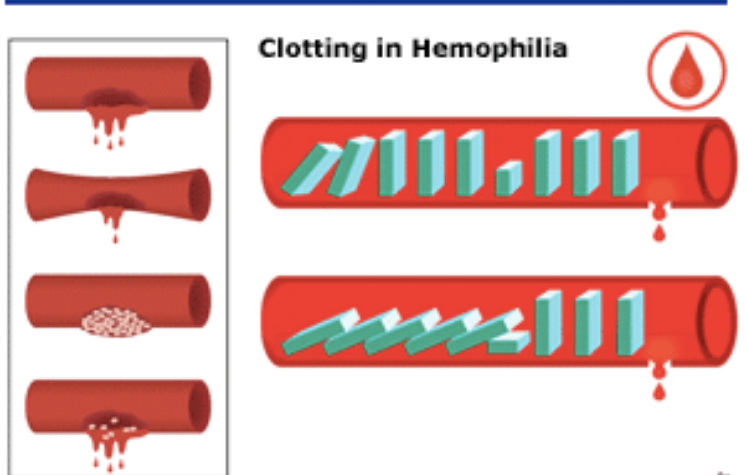
„Minoogowi, rybie bezszczękowej brakuje czynników IX i V . Są to ważne elementy kaskady krzepnięcia krwi, bez których mogą się obejść. Kiedy okazało się, że wielorybom i rybom kostnoszkieletowym brakuje wewnątrzpochodnego układu krzepnięcia krwi, Michael Behe stanowczo stwierdził: „ Tym gorzej dla nich ”. U ludzi brak czynnika IX powoduje zaburzenie krzepnięcia zwane hemofilią B , brak czynnika V powoduje również hemofilię, a brak obu z nich skutkuje ciężkom skazą krwotoczną.

Ryby kostnoszkieletowe i bezszczękowe mają zredukowany zestaw czynników krzepnięcia krwi w porównaniu do ssaków. Rybom bezszczękowym brakuje czynników IX i V. Czynnik tkankowy i czynnik VII bezpośrednio aktywują trombinę. Kaskada krzepnięcia jest inicjowana przez współdziałanie czynników IX i V.
Konsekwencje tego faktu dla „nieredukowalnej złożoności” są daleko idące.”
https://themedicalbiochemistrypage.org/hemostasis-biochemistry-of-blood-coagulation/


https://blog.emergingscholars.org/2014/03/is-evolution-true-the-case-of-intelligent-design/



Russell Doolittle dużo pisze na temat postulowanych mechanizmów powstawania nowych genów [i zwiększających objętość genomu]. Takimi postulowanymi mechanizmami są duplikacje genów i tasowanie egzonów.
http://www.nauka-a-religia.uz.zgora.pl/images/FAG/2004.t.1/art.07.pdf

https://bazhum.muzhp.pl/media//files/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)-r2005_2006-t2_3/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)-r2005_2006-t2_3-s67-96/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)-r2005_2006-t2_3-s67-96.pdf

[…..] Rozważmy [inny] hipotetyczny układ, w którym białka homologiczne do wszystkich części nieredukowalnie złożonego mechanizmu molekularnego z początku pełniły inne indywidualne funkcje w komórce. Czy nieredukowalny system mógł w takim przypadku zostać złożony z pojedynczych składników, które pierwotnie funkcjonowały osobno – jak proponują niektórzy darwiniści? Niestety, jak pisałem w Darwin’s Black Box, zarysowany powyżej obraz znacznie upraszcza ten problem […..] części układu molekularnego muszą automatycznie odnaleźć siebie nawzajem w komórce. Nie może ich ułożyć pewien inteligentny czynnik [……] Aby odnaleźć się wzajemnie w komórce, oddziałujące ze sobą części muszą mieć powierzchnie ukształtowane tak, żeby bardzo dobrze do siebie pasować [….] Pierwotnie jednak funkcjonujące z osobna składniki nie miałyby komplementarnych powierzchni.

Tak można sobie wyobrazić kaskadę krzepnięcia krwi, jak i każdy układ nieredukowalnie złożony
Tak ewolucję - nie ma modelu teoretycznego, który opisywałby ewolucję nieredukowalnych rdzeni kaskady krzepnięcia krwi!
Wszystkie oddziałujące ze sobą powierzchnie wszystkich składników musiałyby więc zostać dopasowane do siebie, zanim zaczęłyby działać razem. Dopiero wtedy mogłaby pojawić się nowa funkcja złożonego systemu. Dlatego mocno podkreślam, że problem nieredukowalności nie znika, nawet jeśli pojedyncze białka homologiczne do składników układu oddzielnie i pierwotnie pełniły swoje własne funkcje.
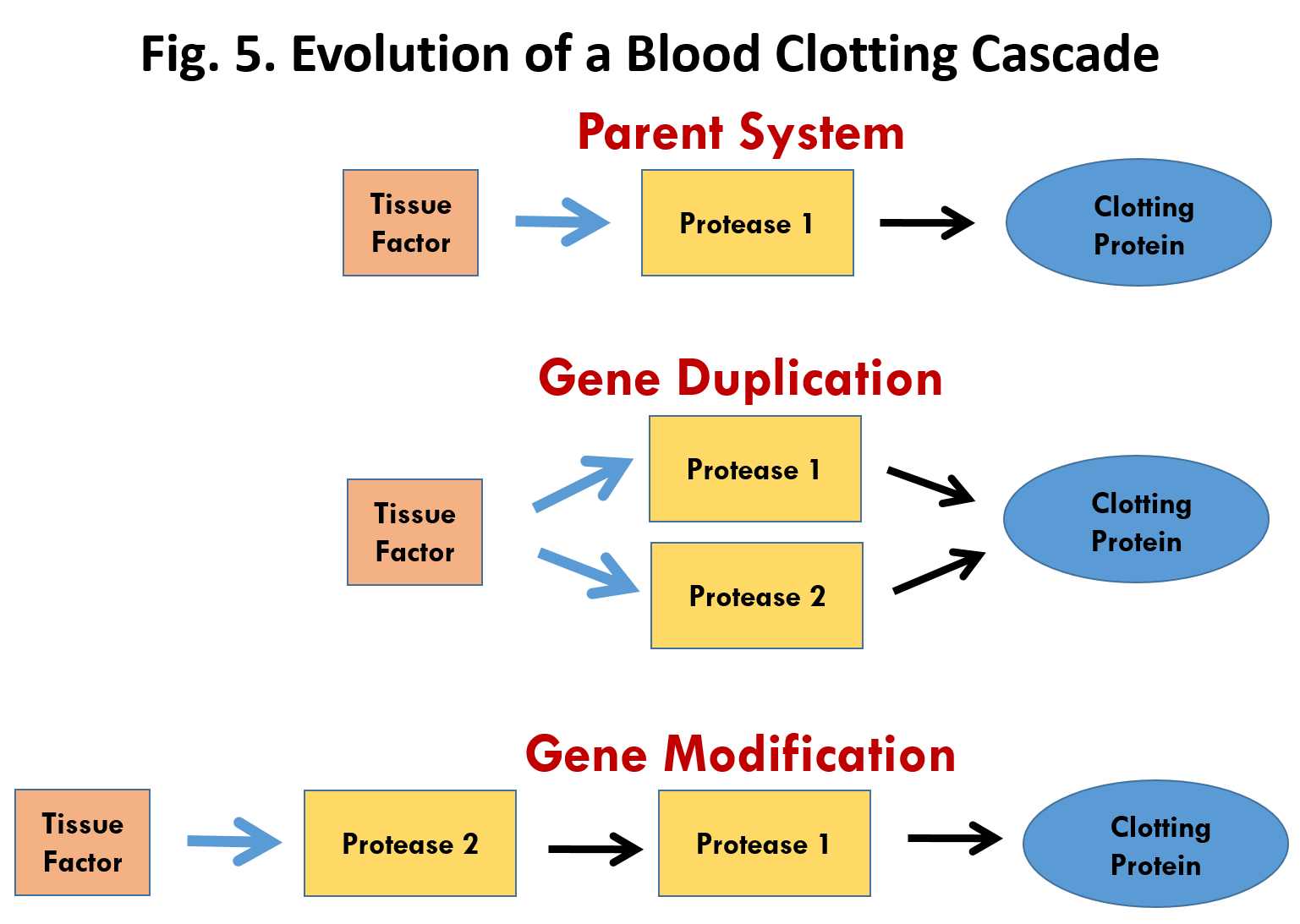
W kaskadzie krzepnięcia krwi jeden składnik wpływa więc na inny, który oddziałuje na następny i tak dalej. Argumentowałem, że kaskada jest nieredukowalnie złożona, ponieważ gdy usunie się jakiś jej składnik, ów proces albo natychmiast się włącza, albo definitywnie wyłącza. Na nic się zdaje – pisałem – postulat, że proces ten rozpoczął się od jednego czynnika, fibrynogenu, po czym dodano inne składniki, gdyż sam fibrynogen do niczego się nie przydaje. Nie warto też zaczynać nawet od czegoś w rodzaju fibrynogenu i niespecyficznego enzymu, który mógłby go rozszczepić, ponieważ krzepnięcie nie byłoby regulowane i możliwe, że czyniłoby więcej szkody niż pożytku.

Tak twierdzę ja. Jednak Russell Doolittle – wybitny biochemik zajmujący się białkami, profesor biochemii w University of California w San Diego, członek National Academy of Sciences, badający przez całe życie system krzepnięcia krwi – nie zgadza się ze mną. Doolittle napisał esej na sympozjum, dotyczący mojej książki i książki Richarda Dawkinsa Wspinaczka na szczyt nieprawdopodobieństwa.
Materiały sympozjum zostały opublikowane wydawanym przez Massachusetts Institute of Technology Boston Review. W eseju tym omawiane jest zjawisko duplikacji genu, dzięki któremu komórka może zaopatrzyć się w dodatkową kopię funkcjonującego genu. Doolittle wysunął przypuszczenie, że składniki procesu krzepnięcia krwi, z których wiele ma strukturę podobną do siebie nawzajem, powstały przez duplikację genu i stopniową dywergencję. Jest to rozpowszechniony pogląd wśród darwinistów (.......) To odwołanie się do duplikacji genu powtórzyło wielu naukowców recenzujących moją książkę, ale odzwierciedla ono powszechne nieporozumienie. Geny o podobnych sekwencjach sugerują tylko wspólne pochodzenie – nie mówią o mechanizmie ewolucji. Jest to istotny punkt mojego argumentu: świadectwo wspólnego pochodzenia nie jest świadectwem doboru naturalnego.(...) Pozwólcie,
że na zakończenie tej sekcji uwyraźnię to, że dwaj bardzo kompetentni naukowcy, profesorowie Miller i Doolittle, obaj wysoce umotywowani w dyskredytowaniu twierdzeń ID oraz gotowi przejrzeć całą biomolekularną literaturę w poszukiwaniu eksperymentalnych kontrprzykładów, obmyślają przykłady, które – gdy spojrzeć na nie sceptycznie – w rzeczywistości raczej wzmacniają niż osłabiają przypadek nieredukowalnej złożoności.
Oczywiście, nie dowodzi to, że twierdzenia nieredukowalnej złożoności są prawdziwe, czy że hipoteza ID jest poprawna, lecz pokazuje – jak sądzę – że naukowcy naprawdę nie wiedzą, jak potraktować nieredukowalną złożoność. Uwidacznia również, że teoria ID jest znacznie mocniejsza, niż uważają jej krytycy.
Wynika stąd także potrzeba traktowania z hermeneutyczną podejrzliwością darwinowskich scenariuszy, takich jakie
zaproponowali Miller i Doolittle. Niektórzy naukowcy tak mocno wierzą w darwinizm, że wydają krytyczne osądy pod wpływem afektów i nieświadomie przeoczają dość oczywiste problemy scenariuszy darwinowskich lub ufnie utrzymują coś, co jest obiektywnie nieprawdziwe( .…) napotkałem gwałtowny sprzeciw – przeważnie ze strony internetowych entuzjastów darwinizmu, którzy pytali, po co w takim razie te setki czy tysiące artykułów badawczych opisujących darwinowską ewolucję nieredukowalnie złożonych systemów biochemicznych. Tworzyli oni strony internetowe, aby owe artykuły udokumentować.
[……]
Nie należy też pochopnie wnioskować o redukowalności systemów posiadających dodatkowe, czyli redundantne składniki, mogą one bowiem posiadać nieredukowalnie złożony rdzeń. Na przykład, samochód z czterema świecami zapłonowymi może jeździć z trzema lub dwiema świecami, lecz z pewnością nie pojedzie bez żadnej. Pułapki na szczury mają często dwie sprężyny w celu zwiększenia ich siły.
Taka pułapka może działać po usunięciu jednej sprężyny, ale nie będzie działała, gdy usunie się dwie.”
http://www.nauka-a-religia.uz.zgora.pl/index.php/pl/component/content/article/38-forum-dyskusyjne/684-russell-f-doolittle-subtelna-rownowaga
„Kaskada krzepnięcia krwi
„Dr Miller wykazał też, że hipoteza o rzekomej nieredukowalnej złożoności kaskady krzepnięcia krwi została obalona eksperymentalnie już w 1969 roku, kiedy stwierdzono, że krew delfinów i wielorybów krzepnie mimo braku pewnej części kaskady, Wyniki te w roku 1998 zostały potwierdzone także na poziomie molekularnym. Niedawno opublikowany został zresztą raport z badań, zgodnie z którym u ryb najeżkokształtnych krew krzepnie pomimo braku nie tylko jednego, lecz trzech składników.(....)”
https://www.evolutionnews.org/2008/12/how_kenneth_miller_used_smokea

Gdyby współczesnemu ssakowi lądowemu usunięto te trzy składniki, którym brakuje rybom bezżuchwowym (znokautowano geny, które je kodują), to czy mógłby przeżyć w przypadku krwotoku – czy dałoby się w ten sposób zredukować ssaczą kaskadę krzepliwości krwi do liczby mniejszej ilości elementów tak, aby dalej była funkcjonalna? Jak to się dzieje, że te ryby, czy inne wymienione przez Millera organizmy z prostszymi kaskadami krwi, przeżywają mimo braku pewnych składników biorących udział w kaskadzie krzepnięcia krwi u ssaków lądowych a te nie?
Od czego to jest zależnie: od biochemicznej organizacji tych różnych systemów - od specyficznego kontekstu molekularnego, czy od środowiska w jakim żyją te różne organizmy? Wniosek z tego taki, że innych rodzajów kaskad nie można traktować, jako FORM PRZEJŚCIOWYCH wiodących do ssaczego typu krzepnięcia krwi. Jedynie czego dowodzą, to wspólnego pochodzenia. Zostały opracowane na bazie podobnych projektów a nie wyewoluowały od wspólnego prekursora w postaci proto-kaskady krzepnięcia krwi.
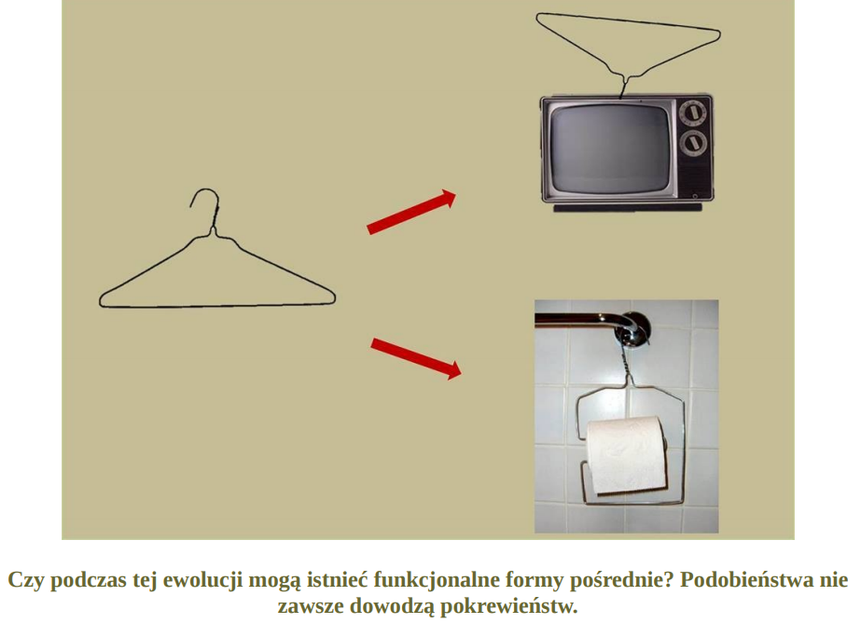

Wyobraź sobie, że do powstania jakieś niezbędnej korzystnej funkcji potrzebne są aż 4 mutacje: A,B,C,D. Że dopiero zestaw A,B,C,D może dać korzyść selekcyjną (dostosowanie). Jaka więc korzyść z pojedynczych mutacji: A, B , C czy D, skoro żadna z nich z osobna nie daje żadnej przewagi selekcyjnej? Każda z osobna jest neutralna, bezużyteczna? W tym przypadku, żeby cecha określana przez mutacje: A,B,C,D mogła dać przewagę selekcyjną, to te 4 [kompletny zestaw] mutacje musiałyby nastąpić za jednym zamachem, a prawdopodobieństwo takiego zdarzenia jest znikome. Jeżeli każda z tych 4 mutacji z osobna nie da przewagi selekcyjnej, to taka ewolucja po prostu nie ma szans nastąpić, ponieważ wtedy nie zadziała dobór kumulatywny.
Istnieje wiele cech, których funkcję określają nierozerwalne zestawy konkretnie ulokowanych w genach nukleotydów. Jeżeli mutacja wprowadzi zmiany w takich konserwatywnych sekwencjach gen przestaje spełniać swoje funkcje i organizm ginie [efekt letalny]. Wiele enzymów, czy białek strukturalnych, posiadają takie konserwatywne domeny (centra aktywne) i jakiekolwiek zaburzenia paraliżują ich funkcję. W podobny sposób zachowuje się enzym zatruty jakimś antybiotykiem. Kontakt z toksyną powoduje, że jego centrum aktywne traci swoją funkcję i przestaje pasować, jak klucz do zamka do konkretnego substratu.
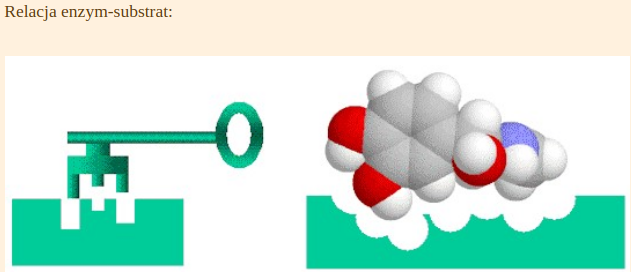
https://botnam.com/enzymes/
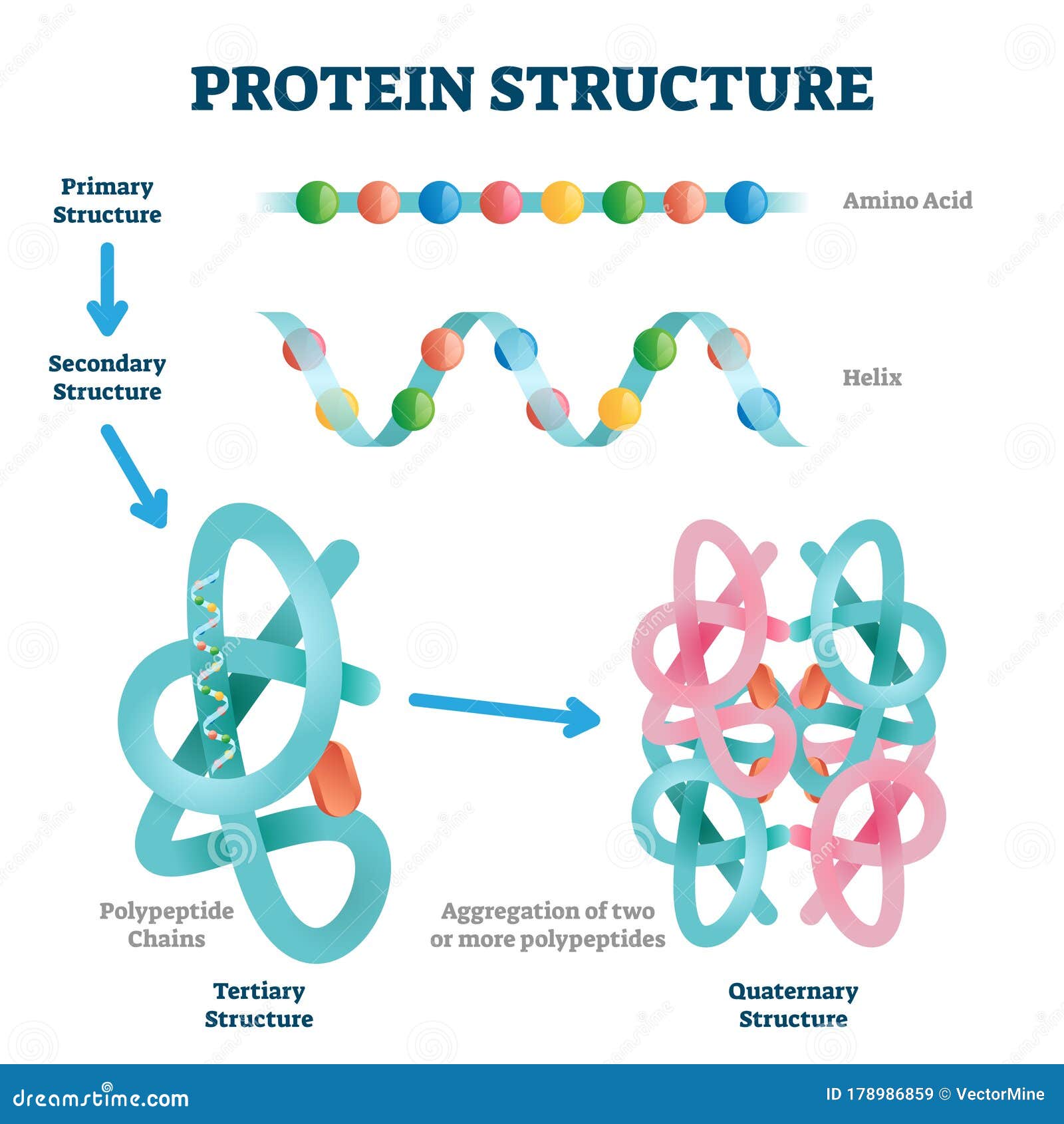
Polipeptyd, to długi łańcuch różnego rodzaju aminokwasów. Jedne z nich nie mają wpływu na formę (konformację, kształt) zwiniętego polipeptydu (białka) inne tak. Rozsiane i ulokowane w konkretnych miejscach wzdłuż łańcucha konkretne aminokwasy odpowiadają za:
-Odpowiednie zwijanie, które następuje według ściśle określonych sekwencji zdarzeń.
-Za powstawanie centrów aktywnych, kieszeni precyzyjnie dopasowanych do danego substratu.
-Za powstawanie komplementarnych domen pozwalających się łączyć z konkretnym typem białka.
https://www.biologyonline.com/dictionary/polypeptide

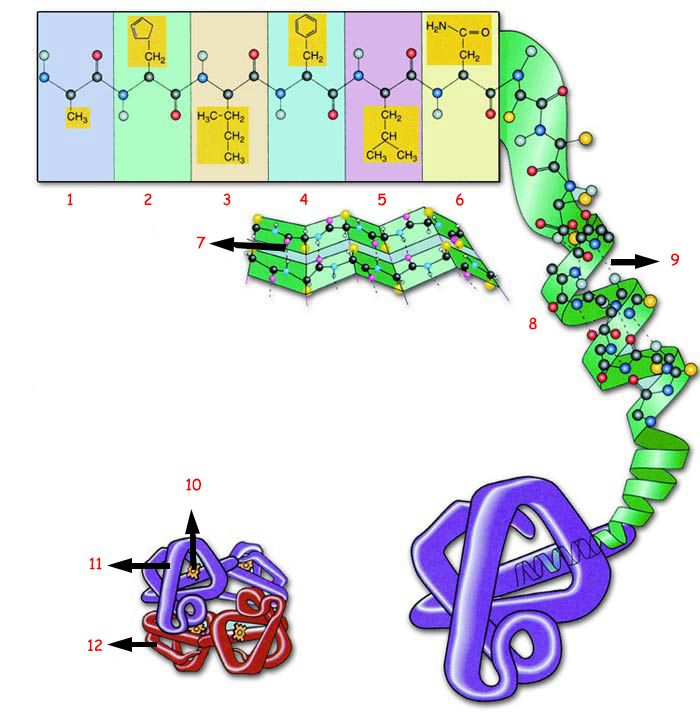
Jeżeli nie pojawiłby się komplet niezbędnych mutacji za jednym razem, tylko powstawałyby stopniowo na przestrzeni długich okresów czasu, to np. białko zwinęłoby się do pewnego momentu i ewolucja utyka w martwym punkcie. Tak też się dzieje w komórce. Zmutowane białka nie zwijają się prawidłowo i dlatego przestają być chronione przez inne białka, zwane chaperonami, ponieważ nie mają się jak przyczepić do źle sfałdowanego białka w celu chronienia go przed zniszczeniem. Wówczas specjalny białko zwane ubikwityną naznacza mutanta do kasacji. Następnie zostaje ono poddane recyklingowi, polegającemu na zmieleniu na poszczególne aminokwasy w specjalnej maszynie zwanej proteasomem:
Proteasom jest to wieloenzymatyczny kompleks utworzony z proteaz. Jest odpowiedzialny za degradację enzymów i białek regulatorowych. Zbudowany jest z cylindra 20S i dwóch regulatorowych kompleksów 19S znajdujących się na obydwu końcach cylindra. Podjednostka 19S rozpoznaje białka, które są przeznaczone do degradacji i odpowiada za jego rozwinięcie i skierowanie do cylindra. Natomiast cylinder jest odpowiedzialny za ich fragmentację. Proteasom degraduje tylko naznaczone wcześniej białka. Znacznikiem tym jest ubikwityna. Wchłonięte przez proteasom białko rozkładane jest do pojedynczych aminokwasów i krótkich peptydów złożonych z 10-12 aminokwasów.
https://www.researchgate.net/figure/The-life-cycle-of-a-polypeptide-inside-a-cell-Modified-from-16-17-reproduced-with_fig1_8009264

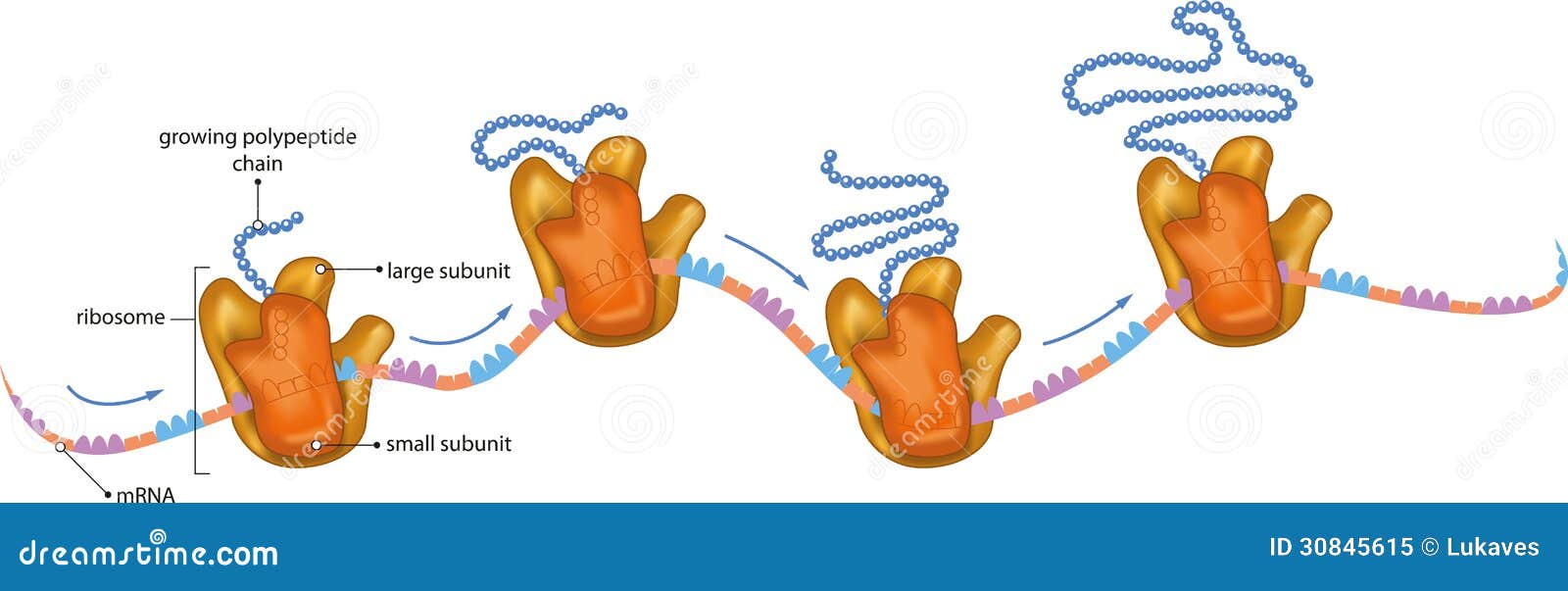
„Cykl życiowy polipeptydu wewnątrz komórki. Po syntezie polipeptydu na rybosomie. Polipeptydy fałdują się w swoją unikalną trójwymiarową strukturę, obejmującą w niektórych przypadkach częściowo zwinięte związki pośrednie.
Te sekwencje pośrednie mają skłonność do agregacji z innymi białkami, jednak proces ten jest niekorzystny kinetycznie, co z kolei sprzyja tworzeniu się odkładających się i kumulujących zespołów toksycznych, które leżą u podstaw takich zaburzeń, jak choroba Alzheimera lub gąbczaste encefalopatie.
W celu wspierania prawidłowego fałdowania łańcucha polipeptydowego w jego pożądaną natywną strukturę, komórka jest wyposażona w zbiór asystentów fałdowania – chaperony, takie, jak białka szoku cieplnego (Hsp) lub białka opiekuńcze.
Białka niezdolne do osiągnięcia pożądanej konformacji trójwymiarowej, zużyte, lub nieprawidłowo sfałdowane są aktywnie degradowane przez komórkową maszynerię kontroli jakości, która obejmuje układ ubikwityna-proteasom (UPS).
Awarie w systemie proteasomów są powiązane z kilkoma zaburzeniami zwyrodnieniowymi. Wiadomo, że rybosomy pomagają we wczesnych stadiach fałdowania białek, co w praktyce oznacza, że bardzo niewiele sekwencji doświadcza procesu fałdowania ze stanu w pełni zdenaturowanego po syntezie w komórce – wiele polipeptydów fałduje się z częściowo zwiniętego.”

Niezbędne są też odpowiednie domeny komplementarnie, jak w przypadku puzzli, dopasowujące odpowiednie białka do siebie. Białka wchodzące w skład kaskady krzepnięcia to głównie proteolizy. Odpowiednie sekwencje pozwalające im ciąć jak piła inne białka byłyby bardzo niebezpieczne, gdyby nieodpowiednie dopasowanie domen pozwalało mieć kontakt sekwencji, którą można porównać do piły mechanicznej, z innymi białkami. Podobne sekwencje mają enzymy trawienne – każdy wie co się dzieje z białkami, kiedy dostaną się do żołądków. Podobnie wyglądałoby środowisko związane z układem krwionośnym, gdyby nie te subtelne dopasowania! Znając te fakty ogólnikowe twierdzenia, że kaskada wyewoluowała z układu trawiennego między bajki można włożyć.

Michael Behe opublikował piękny i madry artykuł, w którym dokładnie wyłożył w czym problem. Zatem darwiniści nie mają jedynie problemów z nieredukowalną złożonością na poziomie systemów biochemicznych, ale też na poziomie białek!
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2286568/






https://slawekp7.files.wordpress.com/2015/01/kaskada-krzepnic499cia-krwi-a-koncepcja-nieredukowalnego-rdzenia.pdf




https://www.researchgate.net/figure/The-schematic-diagram-of-principal-processes-involving-creation-of-fibrins-Prothrombin_fig1_237808077

Schemat poglądowy głównych procesów tworzenia fibryny. Protrombina i trombina to białka aktywujące łańcuch reakcji chemicznych, które aktywują inne białka – fibrynogen. Monomery fibrynogenu tworzą nici fibrynowe.

https://www.researchgate.net/figure/A-Schematic-representation-of-prothrombin-with-its-constituent-Gla-domain-kringles-K1_fig1_332452540

(A) Schematyczne przedstawienie protrombiny z jej składową domeną Gla, domeną kringle (K1 i K2) i domeną proteazy zawierającą łańcuch A i B. Miejsca aktywacji, R271 i R320, są zaznaczone na czerwono. Miejsca ataku proteolitycznego przez chymotrypsynę oznaczono na czarno. Rentgenowskie struktury krystaliczne zamkniętej (B, PDB ID: 6C2W) i otwartej (C, PDB ID: 5EDM) form protrombiny, które ulegają wzajemnej konwersji, wykorzystując elastyczność regionów łącznikowych, takich jak Lnk2. Forma otwarta jest atakowana przez chymotrypsynę w W468 w pętli autolizy. Zamknięta forma jest chroniona przed tym rozszczepieniem przez zapadnięcie się Y93 w kringle-1 do miejsca aktywnego, gdzie angażuje W468 i W547 w interakcjach układania. (D) Schematyczne przedstawienie czterech związków pośrednich konwersji protrombiny (ProT) do trombiny (T). Konwersja obejmuje rozszczepienie przy R271 i R320. Wcześniejsze rozszczepienie zrzuca pomocniczą domenę Gla i domeny kringle i generuje nieaktywną pośrednią pretrombinę-2 (Pre2). To ostatnie rozszczepienie oddziela łańcuchy A i B, które pozostają połączone wiązaniem dwusiarczkowym C293-C439, i generuje aktywną pośrednią meizotrombinę (MzT).


http://zsim.wszia.edu.pl/fizjoterapia/files/2012/01/zsim-34-bsokolowska.pdf




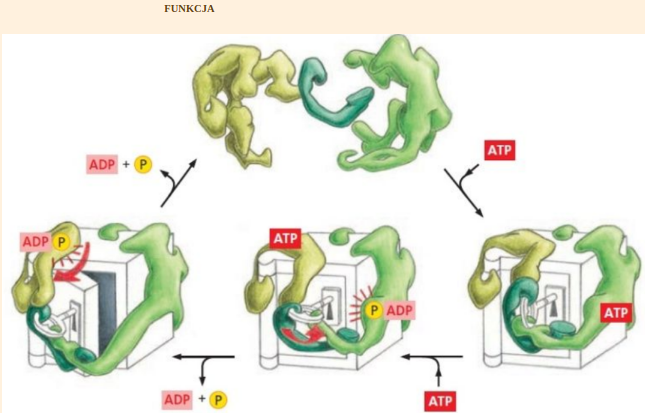

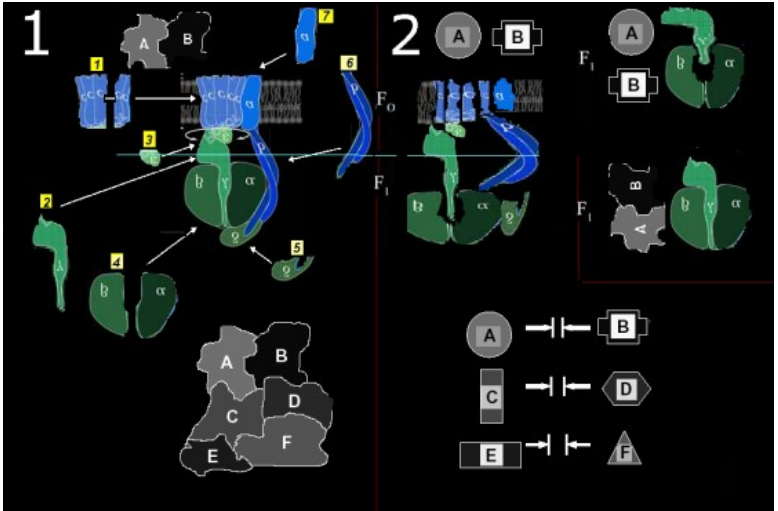
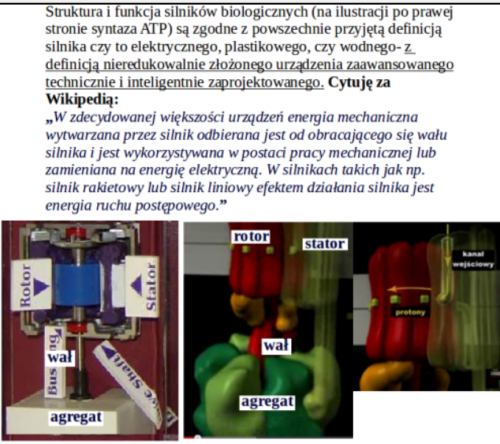
https://slawekp7.wordpress.com/2021/07/28/10108/













https://udel.edu/~mcdonald/mousetrap.html



https://www.researchgate.net/figure/Modeling-of-the-FliF-C-FliG-N-Ring-with-C-25-Symmetry-A-Fully-assembled-FliF-C-FliG_fig8_312399615

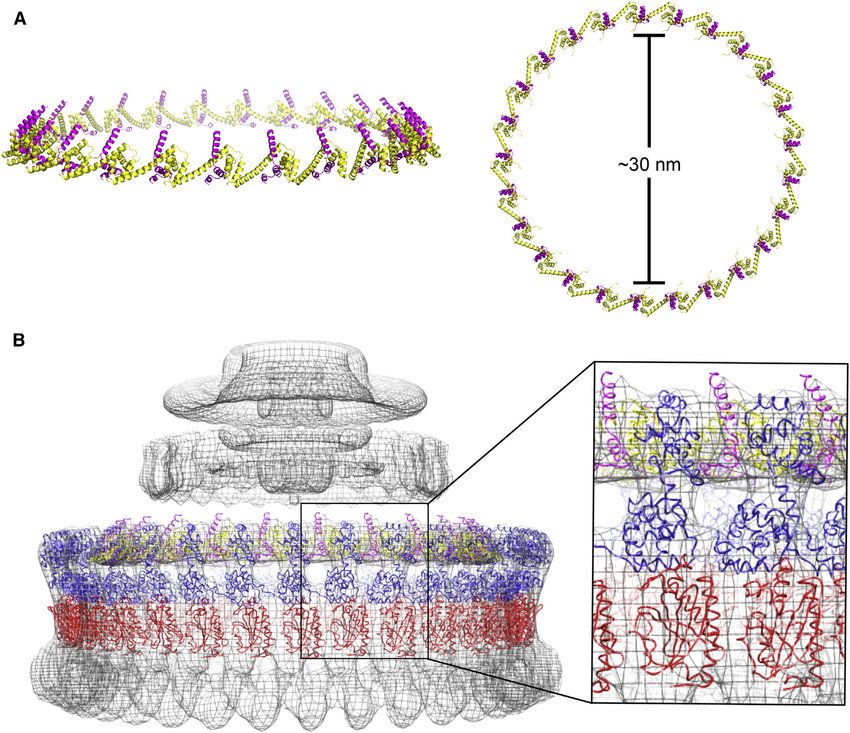
Ważny Wstęp!
ZARYS PODSTAWOWYCH MECHANIZMÓW EWOLUCJI W MODELOWYM KONTEKŚCIE EKOLOGICZNYM
Zwolennicy neodarwinowskiej wizji ewolucji oraz inteligentnego projektu w biologii posługują się tym samym aparatem poznawczym. O tym czym jest założenie (hipoteza), a czym teoria naukowa. Kim są biolodzy doświadczalni i jakie zadania stawiają przed sobą biolodzy teoretyczni. Czym jest funkcja a czym jej geneza
Mało kto ze zwolenników teorii inteligentnego projektu w przyrodzie, nawet z pośród naukowców zgromadzonych przy Discovery Institute, potrafi testować założenia (hipotezy) darwinistów, przy zastosowaniu jedynie dostępnego na rynku naukowym neodarwinowskiego programu badawczego. Żeby to umieć robić należy dobrze rozumieć zakładane mechanizmy neodarwinowskiej ewolucji - w wyniku losowych mutacji korzystnych [np. duplikacji i punktowych] selekcjonowanych przez dobór naturalny:
https://zpe.gov.pl/a/zmiennosc-organizmow/D1B3tf0YZ
https://pl.wikipedia.org/wiki/Dob%C3%B3r_naturalny
Biolodzy doświadczalni badają i poznają funkchję danego procesu biologicznego, natomiast biolodzy teoretyczni (czy jak kto woli: ewolucyjni) po zapoznaniu się z wynikami badań tych pierwszych usiłują zrozumieć ewolucyjną genezę tego zjawiska przyrodniczego. Następnie stworzyć model meoretyczny opisujący poszczególne kroki, jakich musiała dokonać ewolucja, żeby dany system biochemiczny mógł zaistnieć. Ich celem jest też wyjaśnienie, jaką funkcję pełniły systemy prekursorowe (formy przejściowe) i dlaczego mogły zdobywać przewagę selekcyjną, jako systemy niekopletne w porównaniu do współczesnych, które miały z nich wyewoluować i które po usunięciu jednego elementu - z grupy białek tworzących nieredukowalnie złożony rdzeń danego kompleksu -tracą zdolność do uczestnictwa w niezbędnych dla przeżycia procesach organicznych. Biolog doświadczalny może się też specjalizować w biologii teoretycznej.
Hipotezy i teorie naukowe o różnym stopniu wiarygodności
https://pl.wikipedia.org/wiki/Hipoteza
https://pl.wikipedia.org/wiki/Teoria_naukowa
https://pl.wikipedia.org/wiki/Rodzaje_twierdze%C5%84_naukowych
Zaczyna się od ustalania stopnia podobieństw między białkami budującymi jakiś kompleks molekularny, następnie zbiór porównanych białek porównuje się z podobnymi białkami występującymi u innych organizmów (analiza filogenetyczna). Podobieństwa (analogie) nie muszą dowodzić pokrewieństw homologicznych (pochodzenia od wspólnego przodka), ale w przypadku prób stworzenia modelu teoretycznego prawdopodobnych dróg ewolucyjnych danego kompleksu biochemicznego, należy postawić założenie dotyczący homologii. Nie wolno z góry wykluczać homologii na korzyść analogii. Wyniki badań mają powierdzić jedną lub drugą opcję.
Bez stawiania założeń i ich testowania nie można przeprowadzić procesu dowodowego, ale założenia (hipotezy), to nie dowody, a nawet nie teorie. Teorię naukową tworzy przynajmniej kilka dobrze potwierdzonych, pasujących do siebie hipotez opisujących genezę jakiegoś zjawiska biologicznego. W zależności od ilości i jakości potwierdzonych założeń mamy słabą, lub dobrą teorię wyjaśniająca. Żeby teorię można było nazywać faktem zbiór zgromadzonych i pozytywnie zweryfikowanych hipotez musi kreślić ostry obraz wyjaśniający.
Proces przypomina szybę pokrytą zmywalną farbą zasłaniającą duży namalowany obraz. Każda zweryfikowana hipoteza to jedno mniejsze, lub większe kółko powstałe w wyniku zmycia farby w określonym obszarze obrazu. Początkowo pozwala nam to widzieć fragmenty dzieła i stawiać słabe teorie lub alternatywne do nich. Nie można jeszcze poskładać wszystkich potrzebnych elementów do kupy, więc ostatecznego, albo najbardziej prawdopodobnego wyjaśnienia nie ma. Stanie się to dopiero wtedy, gdy miejsc ze zmytą farbą będzie tyle, żeby móc przeprowadzić spójną logicznie syntezę zdobytej wiedzy stwierdzając, co jest namalowane na obrazie.
Okazuje się, że namalowana jest maszynka do mielenia mięsa i to twierdzenie jest prawdziwe, ale pozostaje do wyjaśnienia, czy mielenie jest napędzane ręczną korbą czy silnikiem elektrycznym. Żeby to sprawdzić uczeni dalej projektują i prowadzą badania na podstawie stawianych i weryfikowanych hipotez - starając się zmyć farbę z reszty obrazu. Niemniej mamy już do czynienia z teorią i zarazem prawdą, faktem naukowym. Teraz chodzi jedynie o dopracowanie szczegółów, ale nawet jakby się nie udało, to nie zmieni to faktu, że na obrazie jest namalowana maszynka do mielenia mięsa. Zatem teoria naukowa nie musi być kompletna, żeby można było ją traktować jako prawdziwe, rzetelne wyjaśnienie na gruncie nauki. Nikt nie wątpi, że istnieje grawitacja.
Koncepcja naukowa, jako fakt i teoria
Newton sprawdził to doświadczalnie - podrzucił jabłko do góry a siła ciążenia spowodowała, że spadło na ziemię. Niemniej fizycy po dziś dzień nie mają pojęcia, jakie zjawiska kwantowe są odpowiedzialne za istnienie grawitacji. Zatem w obrębie badań nad grawitacją napotykamy twarde fakty i hipotezy - jeżeli w ogóle istnieją warte poważnego rozważenia. Być może nigdy nie dowiemy się, jakie czynniki są odpowiedzialne za działanie grawitacji, ale czy ktoś rozsadny z tego powodu zaneguje jej istnienie? Neuronaukowcy ponad wszelką wątpliwość udowodnili, że mózg jest odpowiedzialny za świadomość, czy to w wyniku myślenia czy przetwarzania sygnałów napływajacych z otoczenia za pośrednictwem narzadów zmysłów. Istnieje jakiś neurobiolog, który potrafi opisać mechanizmy w komórkach nerwowych, których współpraca pozwala ludziom i zwierzętom odczuwać świadome doznania? Komórki nerwowe, neurony czy glejowe, tworzace mózg składają się, jak wszystkie inne rodzaje komórek z łańcuchów biologicznych: DNA, RNA, z cukrów, polipeptydów, i z błon tłuszczowych.
DNA, RNA, cukry i polipeptydy, które zwijają się w różne białka oraz błony komórkowe złożone są z mniejszych elementów. "DNA i RNA są polimerami (w przypadku DNA, często bardzo długimi polimerami), i są zbudowane z monomerów znanych jako nukleotydy. Kiedy połączymy te monomery, powstający łańcuch jest nazywany polinukleotydem (poly- = "wiele"). Każdy nukleotyd jest zbudowany z trzech części: pierścienia zawierającego azot, pięciowęglowego cukru i przynajmniej jednej grupy fosforanowej." [1] Innymi słowy nukleotydy składają się z atomów.
Polipepotydy z odpowiednio połaczonych aminokwasów. Aminokwasy składają się z atomów wodoru, azotu, wegla i tlenu [2] Cukry (węglowodany, sacharydy)w ścisłym znaczeniu są to organiczne związki chemiczne składające się z atomów węgla oraz wodoru i tlenu.[3] Tłuszcze, lipidy tworzace błony komórkowe - składają się z atomów wodoru, węgla i tlenu [4] Podsumowując tą wiedzę rozumiemy, że mózg składa się z atomów, które tworzą monomeryi polimery, a ich odpowiednia struktura i współpraca pozwala na świadome doznania. Jak to się dzieje, ze ten system uporzadkowanych atomów wywołuje subiektywne wrażenie swiadomości nie wie nikt. Ktoś na podstawie tej niewiedzy zaneguje fakt, ze to mózg jest odpowiedziaklny za swiadomość? Czy ta luka w wiedzy przeszkadza w dalszym badaniu i poznawaniu funkcji mózgu? [5] Istnieją hipotezy, ze za swiadomość odpowiadają zjawiska kwantowe. Jeżeli to prawda, to nikt ich nie zbadał i nie potrafi opisać. [6]
Biologia teoretyczna (ewolucyjna). Narzędzia badawcze i metody dochodzen do prawdy - logika odkryć naukowych [7]
Proces przypomina powolne wznoszenie się podnośnika z obserwatorem. Najpierw widzi niewiele, ale niejako z lotu ptaka coraz więcej, aż w końcu granice jakiejś osady. Dopiero teraz może wykonywać liczne zdjęcia terenu i na ich podstawie kreslić wiarygodne mapy. Dochodzenie do poznania obiektywnej prawdy naukowej można porównać do pracy detektywa, który szuka i co jakiś czas znajduje poszczególne puzle, a następnie stara się je połączyć ze znalezionymi wczesniej. Początkowo różne puzle dają fragmentaryczny obraz, ale z czasem niektore z nich zaczynają do siebie pasować i w tych miejscach układanka staje się coraz lepiej rozumiana. Wróćmy do metod badawczych, jakie stosują biolodzy teoretyczni.
Po ustaleniu stopnia zakładanych podobieństw między białkami buduje się drzewo filogenetyczne [8], a następnie postuluje mechanizm powstania tych białek od wspólnego białkowego przodka. W tym przypadku stosuje się hipotezę duplikacji genów. [9] Gen kodujący jakieś białko ulega duplikacji (podwojeniu) i w efekcie jedna kopia pełni pierwotną funkchję, druga stopniowo zaczyna się różnić od genu wyjściowego, nabierając zdolności do pełnienia innego zadania. Zakłada się, że podczas różnicowania się tego genu powstają kopie bezużyteczne, które wypadają z gry, ponieważ sa neutralne (nie pełnią funkcji wpływającej na przetrwanie organizmu) i tym samym niezauważalne przez dobór naturalny. Dobór widzi tylko te geny, ktore mają wpływ na fenotyp (organizm), a te, które najskuteczniej przystosowują organizm do środowiska zwyciężają z kopiami, które mają mniejsze możliwości.
Przykład z życia na stepie
Załóżmy, że powstały trzy warianty genu odpowiedzialnego za kolor sierści u dzikich królików. Jedne kodują białe futro, inne czarne a jeszcze inne biało-czarne. Które króliki częsciej będą przeżywać ataki drapieżników na białym, snieżnym podłożu? Białe! Będa wydawały coraz więcej potomstwa i białe dzieci będą coraz liczniejsze, aż wypełnią środowisko. Przełoży się to na redukcję ilości drapieżników, gdyż zdobywanie pokarmu w środowisku, gdzie biały królik jest praktycznie niedostrzegalny jest bardzo utrudnione.
W biologii ewolucyjnej funkcjonuje pojęcie selekcyjnego wymiatania [10] i doboru kumulatywnego [11]. Selekcyjne wymiatanie zachodzi wówczas, gdy mutacja dająca przewagę selekcyjną szybko rozprzestrzenia się w populacji. W tym przypadku ta, która przyczyniła się do powstania u królików białego futra. Białe króliki dosłownie wymiotły konkurujace z nimi o zasoby czarne i czarno-białe. Białe zwierzęta żerujące na terenie pokrytym sniegiem były rzadko dostrzegane przez drapiezniki. W oczy rzucały się czarne i czarno-białe króliki, padając łatwym łupem i zostawiając coraz więcej zasobów białym królikom i ich coraz liczniejszemu potomstwu (o tym samym kolorze siersci). Kiedy się dobrze i regularnie odzywia, to urodzi się dużo zdrowego i silnego potomstwa. Jak się dobrze kamufluje na białym podłożu, to wiekszość przeżyje do wieku rozrodczego (nie straci karmiacej matki, nie skończy w żoładku drapieznika).
Początkowo łatwy łup w postaci czarnych i kolorowych króloików wpływa na wzrost populacji drapieżników, co przyśpiesza zagładę królików, ale po jakimś czasie orły zaczynają konkurować o zdobycz, bo czarnych i czarno-białych królików jest coraz mniej. Przyczynia się to do spadku liczebności drapieżników. W końcu pozostają jedynie te nieliczne, które jako tako wyspecjalizowały się w dostrzeganiu i polowaniu na białe króliki.
Kiedy gen utworzy w populacji więcej niz dwa warianty (kodujace białe, biało-czarne + czarne futro) mówimy o allelach wielokrotnych [12] i polimorfiźmie (wielopostaciowości) w populacji [13]. Geny kodujace różne cechy mogą być sprzężone. Np. geny odpowiedzialne za powstawanie białego futra z genami kodującymi czerwone oczy, dłuższe łapy i krótsze uszy. Biało-czarne i czarne króliki mogą mieć geny kodujace kolor sierści sprzężone z kodującymi inne cechy: zielonymi oczami, krotkimi łapami i długimi uszami. Zatem kiedy białe króliki wyeliminują konkurencje tym samym przyczynią się do redukcji polimorfizmu (róznorodności) w populacji. Przetrwają tylko geny odpowiedzialne za białą sierść i te, które są z nimi sprzężone [14]. Geny sprzężone nie muszą mieć wpływu na przeżywalność białych królików, po prostu przechodzą do przyszłych pokoleń podczepione do genów zwiekszajacych szanse przeżycia i wydania potomstwa.
Zanim nastała długa zima białe króliki występowały w mniejszości, ponieważ okazywały się najłatwiejszym łupem pośród trzech odmian tych zwierzat.Były łatwo dostrzegalne na ciemnym tle. Dopiero zmiana środowiska przyczyniła się do szybkiej frekwencji genów kodujących białe futro [15]. Niemniej teoretycznie kolor oczu nie miał żadnego znaczenia dla doboru naturalnego, to cecha neutralna. Zjawisko zabierania w przyszłe pokolenia genów sprzężonych z dajacymi przewagę selekcyjną określa się w biologii ewolucyjnej: genetycznym autostopem [16]. Należy zaznaczyć, ze neutralna cecha, ktora załapała się na genetyczny autostop może dać przewagę selekcyjną w przyszłości.
W przypadku, gdy drapieżnikami były jedynie orły, ratunkiem dla namierzonych przez nie królików było błyskawiczne ukrycie sie w norze. Gdy białe króliki zdobyły dominację, populacja orłów została znacznie uszczuplona. Zaskoczone nagłymi zmianami warunków orły w większości nie zdoły doprowadzić do wieku rozrodczego wymaganej liczby potomstwa. Wiele tych drapieżnych ptaków wyemigrowało w celu szukania bardziej przyjaznych terenów. Niemniej może się zdarzyć, że na teren zamieszkiwany przez białe króliki przywędruje szybko biegający drapieżnik lądowy [17], który żerując na ziemi sprawniej dostrzega ofiarę i skuteczniej poluje. Wówczas białym królikom mogą się przydać dłuższe łapy w porównaniu z wymiecionymi przez selekcyjną kosę pobratymcami, ponieważ umożliwią szybszą ucieczkę.[18]
Dobory jednorazowy i kumulacyjny
W ewolucjoniźmie chemicznym i biologicznym funkcjonują dwa pojęcia mające określać typy doborów naturalnych: dobór jednorazowy i kumulatywny - w obrębie biologii ewolucyjnej dobór naturalny dzieli się na kilka rodzajów: selekcję kierunkową, stabilizującą, na dobór płciowy i krewniaczy.[19]
Dobór jednorazowy działał podczas hipotetycznej ewolucji chemicznej - gdy chemia przekształcała się w biochemię i biologię. W czasie spontanicznego, lecz jak twierdza darwiniści stopniowego, powstawania pierwszej żywej komórki w procesie określanym abiogenezą. Zanim pojawiło się życie komórkowe, zdolne do replikowania materiału genetycznego i rozmnażania dobór naturalny, w tym jego hipotetyczna forma: dobór kumulatywny, nie mógł działać.
Nie było replikacji materiału genetycznego, nie było mutacji i co za tym idzie zmienności. Zatem dobór naturalny nie miał czego selekcjonować, promując proto-replikatory u których powstały korzystne mutacje. Nie ma korzystnych mutacji i konkurencji o zasoby, zdobywania przewagi selekcyjnej przez najsprawniejsze proto-komórki, to i nie ma rozprzestrzeniania się dobrych mutacji w nieistniejacej populacji. A co za tym idzie ewolucji biologicznej, polegajacej na zdobywaniu przestrzeni życiowej i zasobów przez najsprawniejsze replikatory i nastepnie do wygenerowania złożoności życia.
Przykład doboru jednorazowego, to powstanie żywej, zdolnej do rozmnazania komórki za jednym razem! Jednak nawet najprostsze bakterie są bardzo skomplikowane. Można je porównywać do sprawnie działającego i dużego zakładu przemysłowego. Zatem żaden szanujący się darwinista nie przyzna się do takich pogladów. Zwolennicy abiogenezy mają swoje wyjaśnienie: pierwsza żywa komórka musiała powstawać stopniowo - to znaczy za pośrednictwem serii doborów jednorazoiwych.Prace nad ustaleniem genomu minimalnego, polegajace na redukowaniu materiału genetycznego bakterii mycoplasmy genitalis, poprzez systematyczne usuwanie różnych genów przeczą takim poglądom. Badania te pokazują jednoznacznie: redukowanie genomu ma swoje granice, których przekroczyć się nie da. Po osiągnieciu tych granic komórka stanowi nieredukowalny rdzeń, w ktorym oibowiązuje zasada wszystko albo nic! [20]
Fiksacja i kumulacja
O zafiksowaniu się korzystnej mutacji biolodzy ewolucyjni mówią, gdy rozprzestrzeni się w populacji i w niej utrwali. Mówienie o jednej mutacji w jednym genie, to uproszczenie ponieważ niemal wszystkie cechy żywych organizmów są skomplikowane i kodowane przez większą ilość do setek genów. Zmieniają się jedynie warianty genów, jak w przypadku powstawania alleli wielokrotnych, co zaowocowało powstaniem białych, czarno-białych i czarnych królików. Zmiana wariantu genu kodującego dane białko nie zmienia tożsamości tego białka.
Porównanie: na świecie istnieją biali, ciemnoskórzy i czerwonoskórzy ludzie. Czy to znaczy, że od koloru ich skóry zależy, czy będziemy ich nazywali ludźmi? :) Zmutowany gen, który odpowiada za białe umaszczenie królików, w dalszym ciągu jest wyrażany na białko biorące udział w szlaku, który jest odpowiedzialny za syntezę barwnika określającego kolor sierści u królików.
Kto tego nie rozumie, nie rozumie też znaczenia słowa: wariant – wyodrębniona odmiana, wersja czegoś ,jakie funkcjonuje w biologii, gdy mówimy o wariancie, czy wariantach konkretnie zdefiniowanego i sklasyfikowanego genu [21]
Dlaczego pozytywna mutacja teoretycznie utrwala się w populacji? Z tego samego powodu, co hipotetyczne przyczyny zachodzenia doboru kumulatywnego. O tempie w jakim pozytywna mutacja rozprzestrzeni się w populacji, zdominuje ja i się w niej utrwali decyduje pięc podstawowych czynników. Z założenia:
1. Rodzaj presji selekcyjnej związany z konkretnym wyzwaniem środowiska - np. w zależności, czy będzie to dobór kierunkowy, preferujący fenotypy skrajne, odbiegające morfologią od innych przedstawicieli populacji. Czy będzie oddziaływał dobór stabilizujacy, usuwajacy fenotypy skrajne, co rzecz oczywista nie sprzyja transformacjom ewolucyjnym, tylko zastojowi - stazie: https://pl.wikipedia.org/wiki/Ewolucjonizm_(biologia)
2. Tempo mutacji i rodzaj mutacji, co się ma przekładać na tzw.ciśnienie mutacyjne, ktore popycha osobniki z dobrą mutacją do konkurencji, zwycięstw i podboju populacji,
3. Oraz rozmiar populacji. Innymi słowy, czym większa populacja tym większe pole do popisu dla ewolucji na poziomie mutagenezy. Z punktu widzenia neodarwinistów mutacje odpowiedzialne za wszelkiego rodzaju adaptacje, nawet te powstałe w wyniku oddziaływania EWOLUCJI KONTROLOWANEJ przez organizm były losowe. Innymi słowy zdaniem darwinistów skomplikowana i wyrafinowana maszyneria molekularna, złożona z wielu osobnych i nierozerwanie zintegrowanych kompleksów białkowych, które współpracują precyzyjnie się uzupełniając pod względem pełnionych funkcji powstała: na PRZESTRZENI DŁUGICH OKRESÓW CZASU, STOPNIOWO W WYNIKU LOSOWYCH MUTACJI ODCEDZANYCH PRZEZ ŚLEPY, NICZEGO NIE PLANUJACY DOBÓR NATURALNY. Richard Dawkins, który rozpropagował idee zakładające istnienie doboru jednorazowego podczas ewolucji chemicznej i kumulatywnego w czasie ewolucji biologicznej, darując sobie szczegółowe wyjaśnienie istoty drugiego typu doboru, kiedy pisał o tej hipotezie w swojej ksiażce 'Ślepy zegarmistrz - jak ewolucja dowodzi, że swiat nie został zaprojektowany' tak to ujął:
"Wbrew wszelkim pozorom jedynym zegarmistrzem w przyrodzie są ślepe siły fizyczne (...). Dobór naturalny - odkryty przez Darwina ślepy, bezrozumny i automatyczny proces, o którym wiemy dziś, że stanowi wyjaśnienie zarówno istnienia, jak i pozornej celowości wszystkich form życia - działa bez żadnego zamysłu. Nie ma ani rozumu, ani wyobraźni. Nic nie planuje na przyszłość. Nie tworzy wizji, nie przewiduje, nie widzi. Jeśli w ogóle można o nim powiedzieć, że odgrywa w przyrodzie rolę zegarmistrza - to jest to ślepy zegarmistrz." (s. 27)
"Dobór naturalny to ślepy zegarmistrz - ślepy, bo nie patrzy w przód, nie planuje konsekwencji, nie ma celu." (s. 47)
Biorąc pod uwagę interpretacje oxfordskiego profesora biologii Richarda Dawkinsa odpowiednie przypadkowe mutacje selekcjonowane przez ślepy, ale jego zdaniem nieprzypadkowy dobór naturalny, w wyniku długotrwałej i stopniowej ewolucji doprowadziły do WYEWOLUOWANIA NIEPRZYPADKOWEJ EWOLUCJI - opartej na duzym zbiorze rozmaitych procesów molekularnych funkcjonujących w ramach normy reakcji na środowisko! Po pierwsze wyjaśnienia domaga się, to jakie korzyści dawały poszczególne etapy tej ewolucji, ponieważ współczene maszynerie odpowiedzialne za NIELOSOWE adaptacje muszą się składać z kompletnych zestawów genów kodujących białka tworzace te wyrafinowane maszynerie. Jak niekompletne systemy, będace ewolucyjnymi prekursorami współczesnych zdobywały przewagę selekcyjną? Po głebszej analizie tego problemu można dojść do wniosku, że dobór naturalny nie jest ślepy, jak postuluje Richard Dawkins. A nawet, ze musiał mieć charakter proroczy, przewidując że te niekompletne systemy w końcu przekształcą się w maszyny, które w krótkim czasie potrafią dokonać tego, na co przypadkowa ewolucja potrzebowałaby milionów lat! :)
Na jakiej zasadzie powstają adaptacje - na poziomie populacji, czy jednostki?
https://www.jstor.org/stable/10.1086/508960
"Przytoczono dwa mocne argumenty przemawiające za twierdzeniem, że żaden proces selekcji naturalnej nie może odgrywać roli w wyjaśnianiu genezy mechanizmów odpowiedzialnych za adaptację.
Zgodnie z pierwszym argumentem selekcja jest siłą negatywną; może wyjaśniać przyczynę wyeliminowania przez nią konkretnych osobników, ale nie wyjaśni, dlaczego te, które przeżyły (lub ich potomstwo) mają takie cechy, jakie mają (mawia się jedynie, że taka a taka cecha powstała, bo pojawiły się odpowiednie mutacje i presja selekcyjna. Albo, że nie powstała, bo nie pojawiły sie potrzebne mutacje albo odpowiednia preesja selekcyjna).
Drugi argument wskazuje, że explanandum (model opisujący zjawisko w celu wyjaśnienia przyczyn ) i explanans (zbiór faktów mających wyjaśniać istotę zjawiska)są pojęciami na różnych poziomach argumetacji: dobór naturalny jest zjawiskiem działającym na poziomie populacji, podczas gdy adaptacja zachodzi na poziomie indywidualnym.
Tak więc doborem naturalnym można wyjaśnić, dlaczego osobniki w określonej populacji mają określoną cechę, ale nie może wyjaśnić w jaki sposób cecha ta powstała u tej jednostki, zanim sie rozprzestrzeniła i opanowała populację."
Richard Dawkins jest zdeklarowanym neodarwinistą i pomysły, że dobór naturalny działa na całe populacje uważa za naukową herezje. Wszystko zaczyna się przeważnie na poziomie jednostki, u osobnika nosiciela dobrej mutacji. Zmiana musi dawać usprawnienie pozwalające skuteczniej zdobywać pożywienie i inne zasoby w postaci parterów do rozrodu. Potomkowie dziedziczący usprawnienia mają wieksze szanse na własny sukces rozrodczy, ich dzieci i wnuki również. Dopóki u innego członka populacji, który odziedziczył tą dobrą cechę i obecnie wspólnie z innymi dziedzicami tej samej korzystnej mutacji wypełnia środowisko od progu ekologicznego po próg ekologiczny nie pojawi się kolejna mutacja jeszcze bardziej usprawniająca zdolności do zdobywania zasobów, przeżywania i wydawania potomstwa.
Jeżeli szczęście mu dopisze, zdąży spłodzić taką ilość potomstwa, że więcej przeżyje niż umrze w wyniku losowych czynników sprawczych, to skutecznie powieli swoje geny lokując je w synach,córkach i wnukach i wysyłając w odległą przyszłość. Cały sukces, jak i poprzedni, gdy istniały osobniki z jedną dobrą mutacja zostaje osiągnięty na drodze selekcyjnego wymiatania konkurencji - tych nieszczęsników, którym nie dane było posiąść drugiej dobrej mutacji. Podwójnie zmutowane potomstwo w kolejnych pokoleniach opanuje populację i staną się liczni, zamieszkując cały teren od progu ekologicznego, po próg ekologiczny! Jednak radość z sukcesu bedzie trwała do czasu, gdy pojawi się mutant z trzecią dobrą mutację i stopniowa rzeź podwójnych mutantów przez potrójnych mutantów zacznie się od nowa.
Można w ten sposób pisać o poczwórnych mutantach, tych co posiąda piąta i szóstą korzystną mutację - a wszystkie zmiany razem, skumulowane na przestrzeni setek tysięcy i milionów lat prowadzą do nabycia przez potomków złożonej cechy, która zapewni jeszcze większy sukces reprodukcyjny i większą przewagę selekcyjną. Kto pierwszy ten lepszy, a w tych zawodach nie ma zmiłuj się, bo geny są "samolubne [egoistyczne]" i dążą jedynie do skutecznego powielania i tworzenia własnych kopii idąc do celu po trupach! Na takiej mniej więcej zasadzie zdaniem Richarda Dawkinsa i innych neodarwinistów wyewoluowała cała oszałamiajaca złożoność życia ze wszystkimi zapierajacymi dech wyrafinowanymi przystosowaniami, czy to: syntaza ATP, wiciowy silnik bakteryjny, transport pęcherzykowy zależny od klatryny, kaskada krzepnięcia krwi. Twoje rybki w akwarium, kanarek, kot, pies Kowalskiego i ludzki mózg. Jak zapewniają neodarwiniści wszystkie znaki na ziemi, w oceanach i na niebie potwiedzają fakt takiego scenariusza i jedyne zadanie neodarwinistów to konieczność dopracowania szczegółów :)
https://slawekp7.wordpress.com/2021/02/12/gggggggo/
Dobór kumulatywny, a prawdopodobieństwo zajścią ewolucji złożonych cech w szybszym tempie. Mechanizmy umożliwiajace zafiksowanie się [utrwalenie] korzystnej mutacji w populacji
Do pierwszej korzystnej mutacji nie została by dodana druga, do tej trzecia, do trzeciej czwarta i tak aż do szóstej gdyby każda z nich po opanowaniu populacji nie stała na twardym gruncie obecna we wszystkich zwycięzcach. Wiadomo, że z tego czy innego powodu wielu zwycięzców zginęło bezpotomnie, albo zaraz po pierwszym spłodzeniu potomstwa - jeszcze w młodości, w pełni sił i możliwości rozsiewania na lewo i prawo swoich genów. Niemniej przytłaczająca większość przeżywała jedząc, pijąc i płodząc. Zatem straty były szybko rekompensowane, populacja dalejliczna, wypełniajaca wywalczone terytorium od progu po próg ekologiczny - na czterech krańcach stepu! W takim układzie, gdzie zysk netto jest zawsze wiekszy pozytywne mutacje mogą być spokojne - zostały zafiksowane w pionie i poziomie.
Mutacyjny totolotek. Dobór kumulatywny: dwa spojrzenia na istotę tego pojęcia
1 Mało tego, rodzi się dużo dzieci, wnuków, czyli graczy w mutacyjnego totolotka. Jak już przy totolotku, to w jakim kraju istnieje większe prawdopodobieństwo skreślenia sześciu prawidłowych - za jakiś czas wylosowanych liczb? W takim, w którym mieszka pięć milionów ludzi i z tego systematycznie gra milion obywateli, czy w takim gdzie mieszka czterdzieści milionów i gra dwadzieścia milionów? Dokładnie na tych zasadach rozmiar populacji decyduje o tempie ewolucji - im więcej graczy, tym wieksza szansa,że któryś urodzi się z dobrą mutacją z dużym kopem windującym na kolejny szczebel drabiny ewolucyjnej. Częste powstawanie kolejnych wymaganych mutacji szybciej uruchamia proces rozprzestrzeniania się ich na drodze selekcyjnego wymiatania, kumulowania potrzebnych zmian i w efekcie osiagnięcia celu do którego tak naprawdę nikt świadomie nie dążył, np. wyewoluowania mózgów Karola Darwina czy Alberta Ajsztajna.
2) Całą sytuację można porównać do gracza, który skreślił prawidłową liczbe z sześciu koniecznych, nastepnie skreślił tą sama liczbę na dwudziestu milionach kuponów, rozdał je dwudziestu milionom graczy i poprosił, żeby każdy z nich sprobował trafić drugą niezbędną liczbe. Następnie pozbierał kupony z drugą prawidłowo skreślona liczbą. Na dwudziestu milionach kuponach skreślił te dwie trafne liczby, rozdał je dwudziestu milionom graczy i poprosił, żeby probowali skreslić trzecią konieczną liczbe....Proces się powtarza do momentu skreślenia wszystkich sześciu liczb.
Geny i selekcja. Planetoida kontra dinozaury
Usprawnione geny dobrze się powielały, bo okazały się dobrymi genami wypromowanymi przez, z jakiegoś powodu zdaniem Richarda Dawkinsa, nielosowy dobór naturalny - np. nieprzypadkowe uderzenie planetoidy, która unicestwiła dinozaury. Przelatywała akurat bliżej ziemi niż księżyca, a znany jest fakt, że wieksze ciała niebieskie silniej przyciągają planetoidy niż mniejsze ciała niebieskie. Zatem wniosek ten opiera się w zasadzie na nie działających w sposób losowy fundamentalnych prawach rządzacych wszechświatem, a nie na zwykłym mędrkowaniu.
Otwartym tematem jest zagadnienie, czy można uznać za zdarzenie przypadkowe to, że akurat wymarły wszystkie dinozaury i inne takie, a inne organizmy przeżyły - jakieś hadrozaury wprawdzie przeżyły jeszcze milion lat w dobrobycie po tej katastrofie. Musiały dobrze jeść i pić, bo w czasie miliona lat powtawało dużo pokoleń hadrozaurów, ale przecież wyjatek nie może potwierdzać reguły!
https://naukawpolsce.pap.pl/aktualnosci/news%2C379527%2Cniektore-dinozaury-przezyly-wielkie-wymieranie.html
Od powstania życia niestrudzenie płynie rzeka genów. Na przełomie kredy i paleogenu niewątpliwie miała pod prąd i to jak, ale jak widać wszystko dobrze się skończyło i bynajmniej nie wyglada mi to na całkowicie przypadkowe zdarzenie. Poniżej będzie więcej na ten temat!
W pawo, w lewo albo do piachu!
Nie było łatwo, wszystko zawsze mogło pójść w innym kierunku niż poszło i zamiast mózgów Karola Darwina, czy Alberta Ajsztajna powstałaby jakaś forma podobna do ameby, mózg jakiegoś owada, ryby, płaza czy małpy. Wszystko zależało od tego, na którym szczeblu drabiny ewolucyjnej, po której mozolnie wspinała się ewolucja aż wykreowała mózg Darwina czy Ajsztajna i mózgi wszystkich ludzi, coś poszłoby inną drogą, albo nawet trafiło w ślepy zaułek, nie wyewoluowałby mózg Karola Darwina. Skąd byśmy wówczas wiedzieli,jaka jest geneza mózgu Ajsztajna i reszty ludzi? Od początku największego widowiska świata więcej było sposobów na umieranie niż przezywanie. Niemniej rzeka genów płynęła niestrudzenie, nawet pod prąd aż dopłynęła do naszych czasów - ludzi myślących (najczęściej tylko o sobie)!
"Mutacje są losowe, dobór naturalny nie jest proceasem przypadkowym - logiczne uzasadnienie"
Na tle tej całej losowości i kreatywnej bezcelowości ostro zaznacza się architekt złożoności życia - wpradzie ślepy i bezrozumny, ale za to nielosowy, to znaczy nie działajacy w sposób przypadkowy dobór naturalny. Kto twierdzi, że jest procesem losowym, to nie wie co mówi i przez taką kreacjonistyczną propagandę, czy szerzoną przez inną pseudonaukową zarazę, oszukani ludzie wierzą, że ewolucja jest proceswm całkowicie opartym na zdarzeniach losowych. Nic bardziej błednego! Zamiast szerzyć dezinformacje w sieci, mamiąc prostaczków pomyśl.
Jeżeli po ziemi chodzą małe żyjątka i niezmiernie ciężkie olbrzymy, to kogo porwie czynnik selekcyjny: silna wichura? Małe żyjątka, bo jest wiadome że wichura przeważnie porywa najprędzej tych, co ważą mnie. Podobnie, jak w przypadku wspomnianych wcześniej ciał niebieskich o wiekszej w stosunku do mniejszych obiektow kosmicznych masie, przyciągających planetoidy. Zatem w takim układzie jest bardzo prawdopodobne, wręcz pewne, że olbrzymy się rozmnożą, bo nikt nie zna tak silnej wichury, która by je udźwignęła i cisnęła w morze - w zamian tego staną się liczni i będą zamieszkiwać od morza do morza.
Wnioskowanie jest oparty na twardych zasadach logiki naukowej i dobrze zbadanych praw ewolucji. Od początku jest do przewidzenia, że nielosowa selekcja w takim układzie zawsze ciśnie w morze konkurencję. Jest to zasada powtarzalna, zatem o przypadku nie może być mowy. Mimo, że mutacje są losowe, dobór naturalny ślepy i bezrozumny i w efekcie ewolucja nie zmierza do zakładanego celu, tylko do jakiego kolwiek, to biorąc pod uwagę wszystkie te fakty i dowody nie można twierdzić, że mózgi Karola Darwina i Alberta Ajsztajna wyewoluowały w wyniku całkiem losowego nieprawdopodobnego procesu, bo takie przekonanie jest sprzeczne z powszechnym ludzkim doświadczeniem!
Hipotetycznie prawdopodobieństwo powstania każdej kolejnej z sześciu omawianych mutacji
Neodarwiniści twierdzą, ze szanse pojawienia się mutacji niekorzystnych (subletalnych, lub letalnych) są wieksze od możliwości powstania mutacji korzystnych, które pozwolą ich nosicielom na zdobycie przewagi w populacji. Ponadto twiedzą, że dajmy na to w mroźnym klimacie szanse na pojawienie się mutacji umożliwiajacych wyewoluowanie grubego futra są takie same, jak pojawienie się mutacji kodujacych futro, które stanowi marną ilolację przed mrozem. Ze wszystkich mutacji, poza neutralnymi i adaptacyjnymi populację oczyszcza dobór naturalny, tak ze na arenie ewolucji pozostają tylko mutacje dajace korzysci, subtelne usprawnienia, ktorych kumulacja PO ZAWROTNYCH OKRESACH CZASU prowadzi do wiekszych, jeszcze bardziej korzystnych, efektow fenotypowych. Do wyewoluowania maszyn przetrwania - sprawnych ciał, opakowań na samolubne geny, które się najskuteczniej replikują i propagują. Sprawne fenotypu postrzega, jako wehikuły odpowiedzialne za transmisje najlepszych genów do przyszłych pokoleń. Zapomina, lub nie che pamietać, ze na poszczególnych etapach ewolucji te wehikuły były niekompletne, wybrakowane i patzrąc oczami neodarwinisty widzimy coraz wieksze buble w miarę cofania się w czasie - szczebel po szczeblu schodzac w doł drabiny ewolucyjnej!
4. Z punktu widzenia neodarwinistów ważne jest też rozmnażanie płciowe, które pozwala potomstwu udoskonalić daną cechę, lub przyśpieszyć ewolucję dziedzicząc ważne cechy za jednym zamachem. Dajmy na to, jeżeli wymagane są bystre oczy i silne szczęki do skuteczniejszego sukcesu reprodukcyjnego i potomek dostanie jedną z nich od matki a drugą od ojca, to zysk będzie podwójny, co się może przełożyć na przewagę selekcyjną wobec innych członków populacji, którzy posiadają bystre oczy, albo silne szczeki, lub nie posiadają żadnej z tych cech fenotypowych.
Pojęcie doboru kumulatywnego można przedstawiać na dwa sposoby:
1. W przypadku systematycznego kumulowania się niezbędnych zmian w populacji, w celu wyewoluowania systemu molekularnego, czy elementu fenotypu, ktory jest na tyle skomplikowany, że nie mógł wyewoluować za jednym zamachem. Do powstania którego nie wystarczy mutacja w jednym genie, ale wymagane jest, żeby na drodze zakładanego procesu duplikacji genów powstało wiele kopii, które poddane obróbce przez korzystne mutacje punktowe zmieniają tożsamość taksonomiczną i zaczynają pełnić zupełnie inne funkcje. W zależności od cechy, jaka ma wyewoluować czas potrzebny na różne modyfikacje i transformacje liczy się w milionach, lub dziesiątkach milionów lat.
-------
Mechanizmy neodarwinowskie, losowe mutacje punktowe i delecje. Jak w czyms pomogą, to przeważnie na co innego zaszkodzą:
Powstawanie białego ubarwienia w wyniku albinizmu, który może powstać na różne sposoby - za pośrednictwem losowych mutacji w wielu genach. W wyniku uszkodzenia mamy do czynienia z nowymi wariantami genów i częsciowa, lub całkowitą utratą funkcji, a nie z powstawaniem nowych genów:
https://pl.wikipedia.org/wiki/Epistaza
"Epistaza (Epistasis, gr. "stojący na") – w genetyce zjawisko oddziaływania produktów ekspresji jednych genów na inne geny niebędące względem nich allelami. Kilka różnych par alleli warunkuje wówczas pojedynczą cechę. Dzieje się tak w przypadku albinizmu, czyli gdy geny warunkują enzymy należące do jednego szlaku metabolicznego."
https://cdnsciencepub.com/doi/full/10.1139/gen-2020-0003
https://pubmed.ncbi.nlm.nih.gov/10094567/
W biologii pojęcie EWOLUCJA ma różne znaczenia:
Nabywanie białej barwy za pośrednictwem mechanizmów molekularnych, które funkcjonują w ramach normy reakcji na środowisko - ewolucji kontrolowanej przez wyspecjalizowane systemy biochemiczne zakodowane w genach. Ten rodzaj adaptacji nie ma nic wspólnego z ewolucja postulowaną przez współczesnych darwinistów, z załozenia opartej na losowym powstawaniu korzystnych mutacji promowanych przez dobór naturalny. Odpowiednie geny odpowiedzialne za dostosowania do zmian w środowisku w ramach ewolucji konrolowanej dziedziczy się od rodziców, a ich aktywność zależy od impulsów ze środowiska, które stymulują ekspresję i aktywność potrzebnych genów. W zależności od potrzeb są one włączane i wyłączane a w populacji utrwalane za pośrenictwem sygnałów epigenetycznych przekazywanych potomstwu. Geny funkcjonujące w ramach normy reakcji na środowisko nie powstają w wyniku losowej duplikacji genów, które z czasem nabywaja nowych funkcji:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6613075/
https://arctic.au.dk/news-and-events/news/show/artikel/from-brown-to-white-evolution-of-the-polar-bear/
"Niedźwiedź polarny wyewoluował od niedźwiedzia brunatnego, zmieniając kolor futra na biały - idealny do wtapiania się w pokryte śniegiem i lodem środowisko. Dzięki nowym danym międzynarodowy zespół badawczy dowodzi, że oddzielenie niedźwiedzia polarnego od brunatnego przebiegało bardzo szybko.
„Porównaliśmy geny niedźwiedzi brunatnych i polarnych i byliśmy zaskoczeni. Przede wszystkim niedźwiedź polarny jako podgatunek ma mniej niż 480 000 lat. Jest więc znacznie młodsza niż wcześniej sądzono” – mówi profesor Rune Dietz z Centrum Badań Arktycznych, Wydział Nauk Biologicznych Uniwersytetu w Aarhus.
Naukowcy są przekonani, że niedźwiedź brunatny migrował na północ podczas cieplejszego okresu klimatycznego, a gdy później nastał okres chłodu, grupa niedźwiedzi brunatnych prawdopodobnie została odizolowana i dlatego zwierzęta zostały zmuszone do szybkiego przystosowania się do nowych warunków.
Naukowcy pokazują również, że niedźwiedź polarny rozwinął zdolności do życia w Arktyce w ciągu zaledwie kilkuset tysięcy lat – a to bardzo szybka ewolucja.
(Impulsy ze środowiska), zmiany w genach (w ilościach kopii danego genu i wzorach ekspresji genów aktywowanych w odpowiedzi na nowe wyzwania otoczenia) i (być może) działania (w jakimś zakresie) doboru naturalny nie tylko doprowadziły do zmiany ubawienia sierści niedźwiedzia polarnego na biały kolor, ale dostosowały do nowej diety. Za te adaptacje odpowiadają zmiany w genach regulujących transport tłuszczu we krwi i rozkład tkanki tłuszczowej w organizmie.
Kiedy naukowcy porównują zestaw genów niedźwiedzi polarnych z ludzkimi, wyraźnie widoczne są zmiany w genie apoB. Jest to gen, który u ssaków koduje najważniejsze białko w LDL (lipoproteina o niskiej gęstości), powszechnie znane jako „zły” cholesterol.
Jest to bardzo ważna adaptacja do życia w rejonach Arktyki, gdzie niedźwiedzie polarne głównie żywią się mięsem bogatym w tłuszcz pozyskiwanym z upolowanych ssaków morskich, przystosowanych do życia w ekstremalnie niskich temperaturach.
Dieta niedźwiedzi polarnych jest zdominowana przez tłuszcze, jednak w ich tętnicach nie ma złogów (blaszki miażdżycowej), dzięki czemu unikają one chorób sercowo-naczyniowych.
„Zmiany w tym genie mogą decydować o tym, że niedźwiedź polarny radzi sobie z wysokimi stężeniami tłuszczu, cholesterolu i cukru we krwi” - wyjaśnia profesor i lekarz weterynarii Christian Sonne z Centrum Badań Arktycznych, Wydziału Bioscience, Uniwersytet w Aarhus."
W ramach uzupełnienia argumentacji zawartej w powyższym streszczeniu, oryginalnie pokaleczonym neodarwinowskimi określeniami, ktore nie maja nic wspólnego z opisywanym zjawiskiem, ale za to potrafią namieszać w głowach ludziom, ktorzy nie maja odpowiedniej wiedzy (w nawiasach zawarłem sprostowania). Nielosowe mutacje w gorących miejscach mutacyjnych, mechanizmy epigenetyczne – ewolucja w super-tempie! O nowej syntezie ewolucyjnej: https://slawekp7.wordpress.com/2021/06/05/nielosowe-mutacje-w-goracych-miejscach-mutacyjnych-mechanizmy-epigenetyczne-ewolucja-w-super-tempie-o-nowej-syntezie-ewolucyjnej/
Tutaj natomiast skomentowałem jeden z wywodów właściciela kanału "Naukowy bełkot", który nie majac pojecia o czym mówi zamienił swój wywód w pseudo-naukowy bełkot! :) Mieszając pojęcia, procesy i postulowane mechanizmy adaptacje w ramach normy reakcji na środowisko zaprezentował, jako pięć dowodów na losową ewolucję w ujęciu tzw. syntetycznej teorii ewolucji (współczesnego neodarwinizmu):
https://slawekp7.wordpress.com/2021/05/19/fototransdukcja-czy-istnieje-darwinowski-przepis-na-ewolucje-oka/
Z uwagi na fakt, że geny współpracują w grupach, co nazywamy epistaza [22] nie zawsze wystarczy jedna, a nawet dwie czy trzy....pozytywne mutacje w danym genie, żeby zauważył go dobór naturalny, pobłogosławił licznym potomstwem, a to potomstwo jeszcze liczniejszym potomstwem, które podczas walki o zasoby i przestrzeń życiowa pokonuje wszelką konkurencję w efekcie stając się panami i paniami rządzacymi stepem. [23] Nie będę się w tym miejscu nad tym rozwodził, zrobiłem to w dalszej części niniejszego tekstu, za pośrednictwem realnych przykładów zaczerpniętych z biologii molekularnej, a nie wyimaginowanego stepu z królikami w trzech odmianach. Uproszczone modele tego rodzaju pomagaja zrozumieć argumentację darwinistów i nauczyć się posługiwania neodarwinowskim programem badawczym, ale nijak się ma do rzeczywistości, gdy rozważamy możliwość ewolucyjnej genezy złożonych systemów molekularnych:
“[…..] Udzielono licznych odpowiedzi Behe’emu – mieli w tym udział
również biochemicy, łącznie ze mną – które odnosiły się do tego, jak naprawdę powinien on przedstawić obecny stan literatury przy uwzględnieniu podanych przez niego konkretnych przykładów.
W rzeczywistości opublikowano próby wyjaśnienia zagadnień, takich jak powstanie wici, układu krzepnięcia krwi czy biochemicznej podstawy procesu widzenia.
Należy przyznać, że przedstawienie tego, w jaki sposób aktualne dane biologii molekularnej sugerują procesy duplikacji genu,[udziału mutacjui punktowych], tasowania domen i eksonów oraz ewolucji dywergentnej [w wyniku doboru kierunkowego i rozprzestrzenienianiu się pozytywnych mutacji w populacji] nie wyjaśnia, jak NIEKOMPLETNE SYSTEMY zyskują przewagę selekcyjną.
Obecnie jesteśmy jednak na etapie gromadzenia wystarczającej ilościdanych na podstawie sekwencji DNA i trójwymiarowych struktur białek, by w niedalekiej przyszłości przeprowadzać liczne testy przypuszczalnych wyjaśnień ewolucyjnych, co do których można by się spodziewać, że nie będą „takimi sobie bajeczkami”"
Bruce H. Weber
Złożoność biochemiczna :
emergencja czy projekt? [24]
Argument to the future - Ten błąd logiczny popełnia się, gdy się twierdzi, że coś w przyszłości uda się wykazać, choć teraz jest to niemożliwe.
Materiały źródłowe dotyczące wstępu zamieściłem na końcu artykułu.
[1]
https://pl.khanacademy.org/science/ap-biology/gene-expression-and-regulation/dna-and-rna-structure/a/nucleic-acids
[2]
https://pl.wikipedia.org/wiki/Aminokwasy
[3]
https://pl.wikipedia.org/wiki/W%C4%99glowodany
[4]
https://pl.wikipedia.org/wiki/T%C5%82uszcze
[5]
https://slawekp7.wordpress.com/2021/03/07/moralnosc-jest-organem-jak-watroba/
[6]
https://fizyka.umk.pl/publications/kmk/03-Neurokognitywna.pdf
[7]
https://pl.wikipedia.org/wiki/Logika_odkrycia_naukowego
[8]
https://pl.wikipedia.org/wiki/Drzewo_filogenetyczne
https://slawekp7.wordpress.com/2012/01/14/drzewo-ewolucyjne-sciete/
[9]
https://www.researchgate.net/publication/297929904_Duplikacja_DNA_-_mechanizm_rozwoju_nowej_funkcjonalnosci_genow_Duplication_of_DNA_-_a_mechanism_for_the_development_of_new_functionality_of_genes
[10]
https://www.informationphilosopher.com/solutions/scientists/dawkins/
https://en.wikipedia.org/wiki/Selective_sweep
[11]
https://www.quora.com/What-is-the-difference-between-natural-selection-in-one-step-and-cumulative-natural-selection-What-is-cumulative-natural-selection-and-how-does-it-work
https://link.springer.com/chapter/10.1007/978-3-319-10840-7_26
https://www.jstor.org/stable/10.1086/508960
https://www.pnas.org/content/110/27/E2460
[12]
https://pl.wikipedia.org/wiki/Allele_wielokrotne
[13]
https://pl.wikipedia.org/wiki/Polimorfizm_(biologia)
[14]
https://pl.wikipedia.org/wiki/Chromosomowa_teoria_dziedziczno%C5%9Bci
https://pl.wikipedia.org/wiki/Nier%C3%B3wnowaga_sprz%C4%99%C5%BCe%C5%84
[15]
https://pl.khanacademy.org/science/ap-biology/natural-selection/hardy-weinberg-equilibrium/a/allele-frequency-the-gene-pool
https://pl.wikipedia.org/wiki/Melanizm_przemys%C5%82owy
[16]
https://en.wikipedia.org/wiki/Genetic_hitchhiking
[17]
https://pl.wikipedia.org/wiki/Efekt_za%C5%82o%C5%BCyciela
[18]
http://stareaneksy.pwn.pl/biologia/1.php?id=1468682
https://portal.uj.edu.pl/documents/19497540/4cc4170a-9c49-41f0-8911-91b60861c145
[19]
https://www.quora.com/Whats-the-difference-between-single-step-and-cumulative-selection
https://slawekp7.wordpress.com/2015/02/09/o-zaskakujacych-wnioskach-neodarwinistow-nieewoluujace-organizmy-popieraja-teorie-ewolucji/
https://slawekp7.wordpress.com/2013/08/02/klopoty-z-pawim-ogonem/
https://slawekp7.wordpress.com/2013/08/02/mit-hipotezy-doboru-krewniaczego-dobor-krewniaczy-a-nieredukowalna-zlozonosc/
[20]
https://slawekp7.wordpress.com/2021/08/09/abiogeneza-najpierw-rna-najpierw-metabolizm-czy-najpierw-bialka/
[21]
https://pl.wikipedia.org/wiki/Wariant
https://docplayer.pl/13170014-Genetyczne-podstawy-umaszczenia.html
file:///home/slawek/Pobrane/super-pp-journal-manager-315.pdf
[22]
https://slawekp7.wordpress.com/2021/07/28/10108/
[23]
https://slawekp7.wordpress.com/2021/02/12/gggggggo/
[24]
https://bazhum.muzhp.pl/media/files/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)-r2005_2006-t2_3/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)-r2005_2006-t2_3-s121-130/Filozoficzne_Aspekty_Genezy_(Philosophical_Aspects_of_Origin)-r2005_2006-t2_3-s121-130.pdf
[26]
https://pl.sci.biologia.narkive.com/68KwORdW/glowne-problemy-ewolucjonizmu
Trochę historii. Dawne wypowiedzi paleontologa Dawida Mazurka i badaczki pochodzenia życia, współpracownicy noblisty Jacka Szostaka Katarzyny Adamali. Oboje byli zaangażowani w tworzenie Serwisu Ewolucyjnego:
https://pl.sci.biologia.narkive.com/68KwORdW/glowne-problemy-ewolucjonizmu
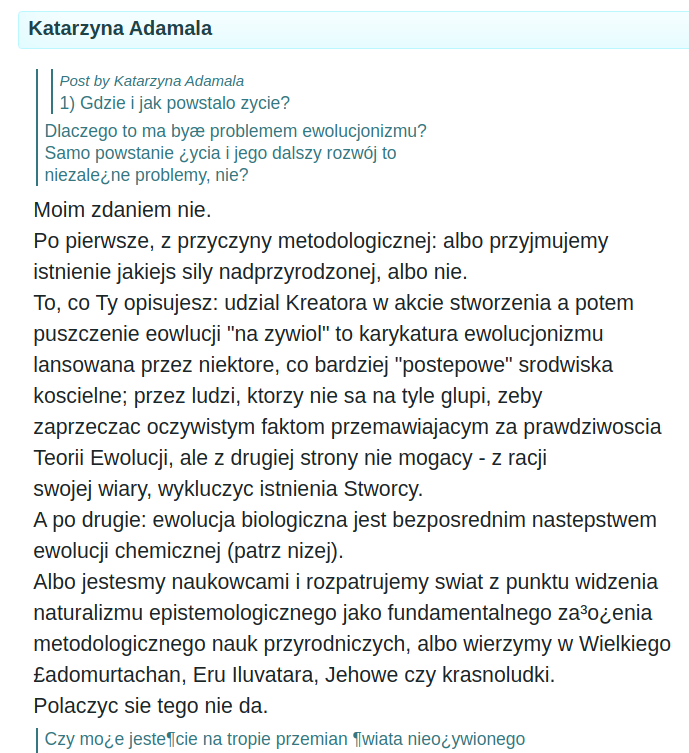

Niestety pani Katarzyna nie zabłysła. W dyskusji brał udział Paweł Poręba i szybko złapał ją na bajkopisarstwie. Tutaj napisałem o Pawle więcej:
https://slawekp7.wordpress.com/2021/05/19/fototransdukcja-czy-istnieje-darwinowski-przepis-na-ewolucje-oka/

Katarzyna Adamala uświadamia sobie, że ma przypał i zaczyna się wycofywać rakiem :)

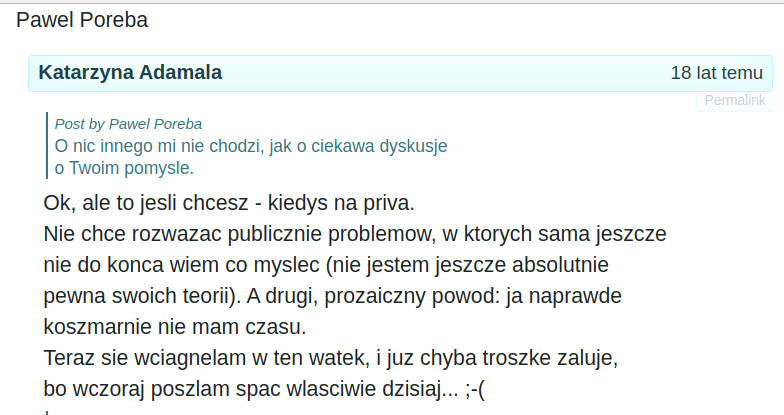
Tutaj można poczytać o współczesnych poczytaniach pani Katarzyny:
https://slawekp7.wordpress.com/2021/08/09/abiogeneza-najpierw-rna-najpierw-metabolizm-czy-najpierw-bialka/
Dawid Mazurek, paleontolog i człowiek, który jeszcze jako student biologii pomagał w rozwijaniu Serwisu Ewolucyjnego:
https://ateista.pl/showthread.php?tid=7898
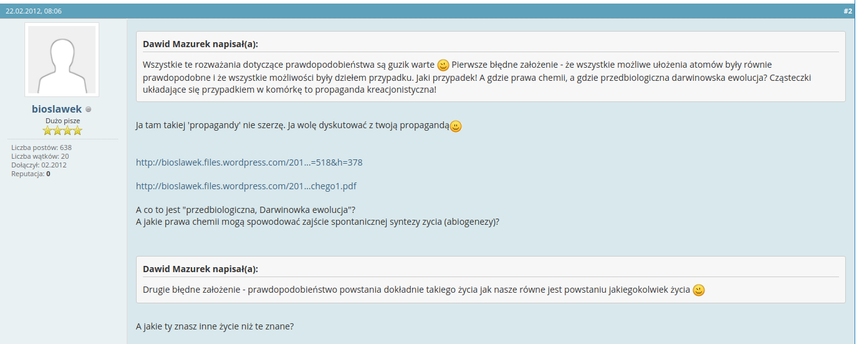
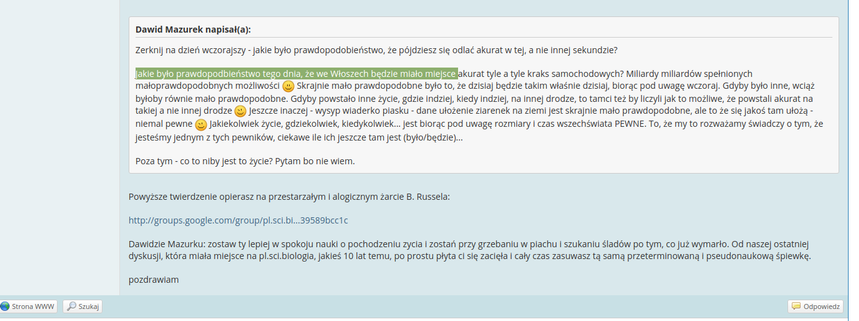
Hahaha... Ten się śmieje, kto się śmieje ostatni!
https://pl.sci.biologia.narkive.com/puR8QtgE/problem-z-protobions#post4
https://groups.google.com/g/pl.sci.biologia/c/rlHw4C6KSFc














No i kto miał rację? Ja dopiero otwierałem przed sobą bramy biologii i biologii teoretycznej, a moi adwersarze byli już magistrami, doktorami i profesorami biologii. Dzisiaj argumentuję niemal identycznie, a oni polegli, lub dosłownie się skompromitowali przed światem uczciwej nauki!
https://www.znaczacy.com/mo-liwo-ci-samoistnego-powstania-ycia-t848270/8



https://pl.sci.filozofia.narkive.com/6dOguCzB/czy-zwolennicy-ip-w-przyrodzie-musz-dowodzi-antytezy

Ta było, jest i będzie, bo prawda naukowa jest jak oliwa i zawsze na wierzch wypływa! :)


Inne tematy w dziale Technologie